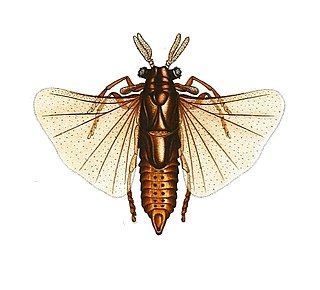
Polistes exclamans
| Polistes exclamans | |
|---|---|
 | |
| Научная классификация | |
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Hymenopterida Отряд: Подотряд: Инфраотряд: Надсемейство: Семейство: Подсемейство: Род: Вид: Polistes exclamans | |
| Международное научное название | |
| Polistes exclamans Viereck, 1906 | |
| Синонимы[1] | |
| |

Polistes exclamans (лат.) — вид общественных бумажных ос рода Polistes семейства Vespidae. Встречается в Северной Америке[1], распространён по всей территории США, Мексики, Багамских островов, Ямайки и некоторых районов Канады[2][3]. Благодаря одиночному основанию гнёзд королевами, P. exclamans расширил свой ареал за последние несколько десятилетий и теперь охватывает восточную половину Соединённых Штатов, а также часть севера. Это расширение обычно связывают с изменением глобального климата и температуры[2].
У ос P. exclamans есть три особые касты, включая самцов, рабочих и королев (яйцекладущих самок), но иерархия доминирования также различается по возрасту. Чем старше оса, тем выше она по рангу в колонии[4]. В большинстве гнёзд P. exclamans есть одна королева, которая откладывает все яйца в колонии. Физиологическое сходство между кастами рабочих и цариц привело к экспериментам, в которых пытались различить характеристики этих двух каст и способы их определения, хотя самцы имеют легко идентифицируемые физиологические характеристики. Поскольку P. exclamans живут в относительно небольших, открытых сотовых гнёздах, они часто подвергаются нападению хищников и паразитов, таких как бабочка-огнёвка Chalcoela iphitalis (Crambidae), наездник Elasmus polistis (Eulophidae) и птиц. У P. exclamans есть стратегии защиты и распознавания, которые помогают защититься от этих хищников и паразитов.
Таксономия
Вид был впервые описан в 1906 году американским энтомологом Генри Лоренцом Виреком[англ.] (1881—1931) по типовому материалу из штата Канзас (США)[5]. Polistes exclamans входит в состав подсемейства полистины (Polistinae) из семейства настоящие осы (Vespidae)[6]. Polistinae (бумажные осы) — второе по величине из шести подсемейств веспид, содержащее около 950 видов, и состоит исключительно из социальных ос. Оно состоит из четырёх триб; P. exclamans входит в трибу Polistini. Род Polistes в настоящее время разделён на четыре подрода, которые распространены по всему миру; P. exclamans входит в подрод Aphanilopterus Нового Света и наиболее близок к P. annularis, P. buysonni, P. canadensis, P. lanio, P. cavapyta, P. simillimus, P. crinitus, P. versicolor и P. instabilis[6].
Описание
В США обитают две формы Polistes exclamans: типичная и изменчивая. Типичная форма встречается на юго-востоке США и обитает в следующих штатах: Северная Каролина, Южная Каролина, Джорджия, Алабама, Флорида, Техас, Луизиана, Оклахома, Арканзас, Канзас, Колорадо и Аризона[3][7]. Хотя окраска этой формы может варьироваться, она почти всегда имеет жёлтую окраску[7]. И проподеум, и мезонотум окрашены в жёлтый цвет. На проподеуме обычно четыре жёлтые полосы, а на мезонотуме — жёлтые линии[7]. На голове также могут быть жёлтые отметины. Крылья, однако, не жёлтого цвета или с жёлтым оттенком, как у некоторых других ос, а инфузорно-пурпурного цвета. Типичную форму часто можно спутать с Polistes fuscatus, Polistes crinitus или Polistes minor[7]. Изменчивая форма имеет красно-коричневую окраску. Середина жгутика, основание брюшных сегментов, внешняя сторона средних и задних голеней, как и крылья, инфускатные или чёрные. У изменчивой формы вместо жёлтых отметин имеются бледные слоново-белые отметины, рассеянные по всему телу[7]. У P. exclamans усики имеют красные, чёрные и жёлтые полосы, в то время как у большинства бумажных ос усики имеют только один цвет. У самок длина передних крыльев может варьироваться от 13,0 до 16,5 мм, а у самцов — от 12,0 до 15,0 мм. Степень выраженности железистых (ржавого цвета) отметин различна[3]. Добычей являются гусеницы бабочек различных семейств, включая таких как медведицы, павлиноглазки, толстоголовки, хохлатки, совки, эребиды, белянки, огнёвки-травянки (Pyraustinae) и бражники[8].
 Вид сбоку
Вид сбоку Личинки
Личинки
Распространение и гнёзда
Встречаются в Северной Америке, распространён в США, Мексике, Багамских островах, Ямайке[1] и в некоторых частях Канады (Онтарио)[2][3]. В США встречается в восточной части страны: Нью-Джерси, Индиана, Иллинойс на юг до Флориды, на запад до Небраски, Колорадо, Оклахомы и Техаса. Также есть единичные находки (случайная интродукция?) в Нью-Мексико, Аризоне и Калифорнии; интродуцирован на Гавайях. Мексика: Чиуауа, Халиско, Идальго[8].
Гнёзда P. exclamans, как у ос-веспид, обычно состоят из бумаги с одним слоем ячеек с открытыми сотами. Размер гнёзда варьируется, верхний предел составляет около 500 ячеек[9]. Хотя многие гнёзда ос имеют внешнюю оболочку из бумаги, гнёзда P. exclamans таковой не имеют[9]. Как указывают Страссманн и Оргрен, «гнёзда примерно круглые, с одной смещённой от центра ножкой [базальной частью], обычно расположенной в верхней части гнезда. Ячейки возле ножки являются самыми старыми»[9]. P. exclamans способны колонизировать новые места, поскольку самки-основательницы часто рассеиваются на новые территории[2]. Гнёзда P. exclamans часто находили вблизи искусственных сооружений и они охотнее строили гнёзда в них и рядом с ними. Из шести исследованных видов ос P. exclamans была единственной, которая занимала искусственные места для гнездования. Она предпочитает хорошо освещённые, открытые места[10].

Гнёзда ос-полистов могут быть построены из древесных волокон, которые собирают со столбов и стеблей растений. Волокно формируется в бумагоподобные соты с шестиугольными ячейками. Гнёзда ориентированы вниз и удерживаются одной нитью[11]. P. exclamans также были замечены в искусственных гнёздах, созданных исследователями и состоящих из пучков соломинок[2].
Цикл жизни колонии
Цикл колонии Polistes exclamans длиннее, чем у северных видов ос, короче, чем у тропических, и, таким образом, является «в некоторых отношениях промежуточным» между ними. Их цикл длится с марта по сентябрь[12][13]. Первый выводок рабочих появляется между маем и июлем. Для развития яиц требуется от девяти до четырнадцати дней, хотя яйца в меньших гнёздах и отложенные позже в сезон, развиваются гораздо дольше: от тринадцати до восемнадцати дней для развития личинок, около тринадцати дней для развития куколок, и в общей сложности от шести до восьми недель для превращения яйца во взрослую особь[9]. Репродуктивные осы появляются позже, в августе или сентябре[13].
Королева
Характеристики
Polistes exclamans живёт в иерархическом обществе с одной королевой, самкой которая откладывает яйца. Однако все самки имеют одинаковую морфологию и обладают физиологической способностью брать на себя откладку яиц в случае необходимости (например, из-за смерти или миграции королевы)[12][14]. Поскольку королева имеет ту же морфологию, что и остальные рабочие в колонии[15], возникает вопрос о том, как распределяется роль королевы и что делает её особенной. Исследования показали, что у королев и самцов уровень глюкозы, фруктозы и трегалозы выше, чем у рабочих, что приводит к повышению уровня криопротекторов. P. exclamans используют эти сахара в качестве криопротекторов, которые работают за счёт увеличения концентрации растворителей в клетках. Это приводит к повышению выживаемости в холодную погоду. Было обнаружено, что при температуре 5 °C выживаемость королев составляет 76 % по сравнению с 17 % выживаемости рабочих особей в течение 15-дневного периода испытаний. Королевы также имеют 0,5-миллиметровый слой жира вокруг своего тела. Это позволяет им дольше жить зимой и, возможно, продлить брачный сезон[15]. Королевы, возможно, единственные, кто способен впадать в диапаузу[16].
Развитие и определение
Хотя внешних морфологических различий между королевами и рабочими особями немного, есть некоторые внутренние признаки, по которым можно определить социальный статус осы[16]. Также было обнаружено, что определение касты необратимо зависит от температуры на незрелых стадиях развития. В одном из экспериментов некоторые осы, появившиеся на свет в июне, содержались в камерах длинного дня, то есть в камерах с температурой 26 °C, в которых 16 часов светло и 8 часов темно. Эти особи имели более крупные и активные яичники по сравнению с теми, кто содержался в камерах короткого дня, установленных при температуре 22 °C с 14 часами света и 10 часами темноты[16]. Предполагается, что это явление может происходить из-за секреции ювенильных гормонов, определяющих касты, и подавления яичников рабочих особей под воздействием окружающей среды или гормональных факторов[16].
Геронтократия
Учитывая, что гнёзда Polistes exclamans часто разрушаются или страдают из-за смерти королевы (чаще всего в результате естественных обстоятельств), необходимо и целесообразно, чтобы существовала особая система замены королевы[17]. Существующую систему лучше всего «описать как геронтократию»[17]. Более того, она также тесно связана с системой доминирования в колонии. В иерархии доминирования старшие особи занимают более высокие позиции, в то время как молодые особи занимают более низкие позиции[17]. Эти старшие самки обычно более агрессивны и чаще добывают корм[17]. В отражение этого ранжирования доминирования и поведения самок видно, что когда королева больше не присутствует в колонии, следующая по старшинству самка становится новой королевой[17]. Это может быть связано с генетикой следующей старшей королевы, так как замена королевы влияет на общее родство самок в выводке, тем самым позволяя будущему выводку быть более родственным взрослым рабочим, которые их воспитывают[17].
Роль
У P. exclamans гнёзда, как правило, меньше, чем у других ос: в одном гнезде обычно менее ста особей. Королева, как правило, является самой активной особью в гнезде[18]. Это связано с тем, что королева должна следить за деятельностью гнезда и непосредственно контролировать ее. В дополнение к своей репродуктивной роли королева также должна действовать как лидер гнезда и синхронизировать активность рабочих[18]. Однако эти два последних пункта недавно были оспорены, и рабочие могут быть самоорганизованными[19].
Размножение и развитие
Система определения пола
P. exclamans — гаплодиплоидные насекомые, как и другие виды Polistes, включая Polistes metricus, Polistes dominula и Polistes annularis. Это означает, что у них есть гаплоидные самцы, которые производят идентичные гаплоидные сперматозоиды, и диплоидные самки, которые производят гаплоидные яйца через мейоз. Это влияет на генетическое родство внутри колоний, поскольку сестры получают идентичную сперму, если у них общий отец, а нормальный полуидентичный вклад — от матери. Это приводит к конфликтам между королевой и рабочими в отношении соотношения полов, причём рабочие предпочитают более женское соотношение полов, а королевы — равные инвестиции в королев и самцов[20].
Половые инвестиции
У Polistes exclamans равное соотношение полов достигается, когда только 46,3 % инвестиций посвящено самкам, поскольку самки ос в 1,16 раза крупнее самцов. В исследовании, проведённом Страссманном, было обнаружено, что половые инвестиции предрасположены к самкам, особенно в годы высокого хищничества и когда гнёзда обычно менее успешны[13]. Было замечено, что у Polistes exclamans самки производятся раньше самцов. Этот перекос в сторону женских инвестиций может быть обусловлен способностью самок становиться либо рабочими, либо репродуктивными самками (это более адаптивно для гнезда, поскольку обеспечивает ему большую гибкость по сравнению с самцами и может быть важно для общего успеха гнезда) или тем, что самки обеспечивают колонии большую защиту гнезда. Даже когда прежней королевы больше нет, половые инвестиции все равно перекошены в сторону самок, чего нельзя ожидать, учитывая генетические последствия появления новой королевы[13].
Ранние самцы
Рабочие особи впервые появляются в период с мая по июль. Во время этого первого появления репродуктивные самцы также появляются вместе с первым выводком и называются «ранними самцами». Эти самцы обеспечивают большую адаптацию, поскольку они допускают присутствие других репродуктивных самцов. Гнёзда с большим числом ранних самцов производят больше рабочих, ячеек, куколок и выводков[12]. Хотя причины этого неясны, гибель цариц у Polistes exclamans — обычное явление, чаще всего в мае. Большинство первоначальных королев погибает к июлю, «задолго до того, как будут отложены яйца, ставшие осенними репродуктивными особями», поскольку репродуктивные особи появляются с конца августа по сентябрь[12]. Таким образом, королевы должны оставаться живыми после июня, чтобы «родить» будущих репродуктивных особей, а если они не могут этого сделать, то новой королевой становится самый старший бывший рабочий[17].
Привлечение полов
Как и многие другие насекомые, P. exclamans используют половые феромоны для привлечения представителей противоположного пола. Исследователи попытались определить, какую именно роль играют половые феромоны в сексуальном влечении бумажных ос. В одном из экспериментов они установили аэродинамическую трубу, где самцы и самки подвергались воздействию фитиля с половыми феромонами, выделенными от самцов и самок[21]. Эти феромоны были получены с помощью гексановых экстрактов из целых тел и тораксов не спарившихся самок. Экстракты самцов были взяты из мандибулярных и стернальных желёз. Было обнаружено, что противоположный пол привлекается при подветренной стороне запаха, и результаты усиливаются, если включить вентилятор. Дальность действия запаха составила около 2 метров[21].
В природе было замечено, что самцы отходят от гнезда, чтобы привлечь самок. Самцы прижимают стернит брюшка к ветке и трутся о неё жвалами. Это используется для привлечения самок, и было замечено, что самки посещают эти ветки и пробуют запахи[21].
Поведение и экология
Присутствие расплода и кастовая дифференциация
У Polistes exclamans сравнительно меньшие гнёзда и колонии по сравнению с некоторыми другими эусоциальными видами ос. Поэтому часто гнездо разрушается, королева умирает до конца сезона, или гнездо разрушается другими способами, такими как хищничество, паразитирование или гибель рабочих, что приводит к тому, что рабочие помогают воспитывать расплод, который менее родственны им, чем их сестры осы[11]. В связи с этим было бы выгодно, чтобы существовали осы, которые остаются бескастовыми до взрослого этапа развития, позволяя самкам ос быть пластичными[11]. Таким образом, было предсказано, что рабочие осы P. exclamans будут действовать как рабочие, если присутствует расплод, и начнут развивать характеристики гин, если выводка нет, демонстрируя вышеупомянутую адаптивную пластичность[11]. Экспериментально было установлено, что это так и есть. Солис и Страссман провели исследование, в котором у экспериментальной группы были удалены яйца и личинки. В экспериментальной группе осы стали вести себя как будущие королевы и даже обзавелись характерным слоем жира, свойственным королевам. В контрольной группе осы продолжали вести себя как рабочие. Это свидетельствует о том, что присутствие расплода влияет на дифференциацию каст и что самки P. exclamans проявляют адаптивную репродуктивную пластичность[11].
Распознавание родства
Распознавание соплеменников по гнезду широко распространено у многих эусоциальных насекомых. Однако при отсутствии физических различий в рационе или материалах для гнезда многие из них уже не могут отличить одного сородича из своего гнезда от других представителей своего вида. Однако, было установлено, что многие насекомые обладают способностью различать родственников, даже живя в идентичной среде с неродственником. Исследования показали, что у ос Polistes fuscatus бывшие сородичи следующей весной гнездятся в местах, расположенных недалеко друг от друга, рядом с их родными гнёздами. Социальные пчёлы-галиктиды также способны узнавать своих родственников[22]. У P. exclamans было показано, что дискриминация родственников от неродственников возможна, даже если они живут в идентичной среде. Это было обнаружено Алленом, Шульце Келлманом и Гамбоа в ходе эксперимента, в котором спящие зимующие осы из разных гнёзд были помещены в коробку вместе после воспитания в идентичной среде. По мере увеличения числа неродственных групп ос в коробке образовывалось все больше скоплений. Это указывает на то, что осы были способны отличать родственников от неродственников, что является адаптивной способностью при защите своего гнезда[22].
Зимовка
Многие виды Polistes объединяются на зиму. P. exclamans собираются в довольно большие скопления во время спячки[23]. "Эти скопления часто встречаются в защищённых местах, называемых гибернакулами, которые могут быть самыми разнообразными: расщелины и трещины скал или стволов, под корой деревьев, между стенами зданий или любые другие естественные или искусственные сооружения, обеспечивающие защиту во время спячки. "Хотя P. exclamans были найдены в гнёздах осы Trypoxylon politum (род Trypoxylon) — грязевом гнезде, которое может обеспечить определённый уровень защиты, не считается, что в нём действительно зимуют P. exclamans[23].
Смертность рабочих особей
Типичный рабочий P. exclamans живёт от 14 до 16 дней. Возраст самой старой осы, наблюдавшейся в природной колонии, составлял 102 дня. Королевы обычно живут в 6 раз дольше рабочих[4]. Наблюдаются некоторые различия в продолжительности жизни P. exclamans. Как правило, это связано с колонией происхождения. Хотя между колониями существуют различия, между исходным гнездом и гнездом-спутником они, как правило, меньше[4]. Различия внутри одной колонии могут также происходить из-за наличия разных каст[24]. Фуражиры, те, кто часто путешествует за пределами гнезда, ведут более рискованный образ жизни и, как правило, имеют более низкую продолжительность жизни, чем те, кто остаётся в гнезде. Тем не менее, было замечено, что колонии с высоким уровнем фуражирования также имели более высокий уровень репродуктивности, чтобы компенсировать более низкую продолжительность жизни[4]. Другой аспект, наблюдавшийся в одном гнезде, заключался в том, что увеличение численности самок привело к снижению продолжительности жизни. Это объясняется тем, что самки были больше сосредоточены на борьбе за звание следующей королевы, чем на добыче корма и выполнении своих функций[4].
Гибель рабочих является важным фактором, приводящим к гибели колонии, и является причиной 13—76 % случаев гибели колоний. Отчасти это может быть связано с малым размером колонии. Поскольку колонии маленькие, продолжительность жизни рабочих имеет решающее значение для выживания гнезда[4].
Гнёзда-спутники

Гнёзда-спутники — обычное явление для P. exclamans. Королева отлетает на расстояние от 0,15 до 11 м от первоначального гнезда, чтобы обосноваться на новом месте. Около 16—39 % гнёзд создают спутники с мая по июль[12].
Подобно осам вида Parischnogaster alternata, которые строят множество гнёзд в кластерах, чтобы создать защиту за счёт эффекта разбавления, спутники используются как страховка от нападения хищников и паразитоидов[25]. Из 12 гнёзд, в которых были спутники и которые были разрушены птицами, 66,7 % колонии выжили, перебравшись в гнездо-спутник, по сравнению с более низким репродуктивным успехом гнёзд без спутников (5,7 %). При вторжении бабочки-паразитоида Chalcoela iphitalis наличие спутников не повысило выживаемость, хотя и оказало влияние при внедрении Elasmus polistis. Это дало P. exclamans шанс на выживание[12].
Рабочие, которые старше, обычно основывают гнёзда-спутники и имеют более развитые яичники. Более молодые и менее развитые рабочие присоединяются к спутнику после его создания. Распределение рабочих между основным гнездом и спутником очень важно, поскольку они необходимы для создания спутника и для продолжения работы основного гнезда, хотя через несколько месяцев основное гнездо может быть заброшено. Если недостаточно рабочих последует за первым рабочим в новое гнездо, то оно, скорее всего, окажется неудачным как спутник[12].
Взаимодействие с другими видами
Паразитоиды
Polistes exclamans живут в общественных гнёздах с открытыми сотами, что делает их доступными некоторым другим организмам. Это делает ос очень восприимчивыми к атакам хищников и паразитоидов. Два наиболее распространённых паразитоида — это молевидная бабочка Chalcoela iphitalis из семейства огнёвки-травянки (Pyralidae) и наездники родов Elasmus (Elasmus polistis, Eulophidae) и Pachysomoides (Ichneumonidae) из отряда перепончатокрылых[26].

 Самец Elasmus
Самец Elasmus
C. iphitalis и E. polistis
Осы P. exclamans предпринимают несколько контрмер против вторжения. Самки Chalcoela iphitalis откладывают яйца в осиные гнёзда по ночам, хотя их часто видят и днём, сидящими на ветке рядом с осиным гнездом. Когда самка C. iphitalis проходит рядом с гнездом, а оса пересекает её след, оса реагирует на это яростно, многократно жаля и кусая то место, где находилась бабочка. Другие осы настораживаются, вероятно, из-за вибрации гнезда, и начинают двигаться по гнезду, начнут двигаться рывками, исполняя характерный «танец паразита» («parasite dance»). Осы покидают гнездо и ходят по всему окрестному субстрату. Если бабочка найдена (что случается редко), она немедленно уничтожается. Осы продолжают свой танец паразита в течение как минимум 10 ч после обнаружения самки бабочки C. iphitalis в гнезде или рядом с ним. Однако это редко происходит. В итоге бабочка откладывает яйца в гнездо ос, а когда они вылупятся, то займут все гнездо. В результате многие куколки ос прерывают своё существование. Обычно это наблюдается в конце лета, когда моль наиболее многочисленна[26].
Второй распространённый паразитоид, наездник Elasmus polistis, также оказывает катастрофическое воздействие на гнездо ос P. exclamans. Из одной ячейки вылупляется до 80 особей E. polistis. Первыми появляются самцы, которые затем покидают гнездо и ждут появления самок. Как только самки появятся, самцы начнут спариваться и размножаться, быстро уничтожая популяцию P. exclamans[27]. В некоторых случаях захватчики E. polistis прячут личинок в гнезде, чтобы P. exclamans не смогли их найти. Хозяева пытаются удалить как можно больше паразитоидов, чтобы предотвратить их распространение на соседние гнёзда. Эффективность этого защитного механизма не доказана, так как чаще всего личинки E. polistis остаются незамеченными хозяевами. В одном из экспериментов было установлено, что в 1981 году более 60 % всех гнёзд потеряли расплод из-за паразитоидов E. polistis или C. iphitali[27]. Также было замечено, что более крупные колонии подвергаются большему риску паразитирования. Это связано с тем, что крупные гнёзда, как правило, старше[28]. Эту связь между возрастом и паразитизмом можно проследить на примере того, что гнёзда старых цариц, как правило, паразитируются больше, чем гнёзда молодых цариц[4].
Хищники
Есть два типа хищников, которые удаляют расплод из всего осиного гнезда сразу, уничтожая при этом гнездо: птицы и муравьи. Птицы обычно разоряют гнёзда на рассвете или в сумерках, убивая при этом немногих взрослых ос. Хищничество птиц в гнёздах легко обнаружить, поскольку все гнездо исчезает сразу. Его останки часто можно найти под ветками, где птица, предположительно, сидела и поедала выводок. В большинстве наблюдаемых лет птицы уничтожают больше гнёзд, чем любой другой хищник или паразитоид. Пик хищничества птиц на гнёздах приходится на май или июнь, хотя оно присутствует с марта по октябрь. После того как гнездо разрушено, оставшиеся взрослые особи пытаются восстановить гнездо. Обычно эта попытка не удаётся, поскольку взрослые особи, скорее всего, погибнут до того, как в новом гнезде появятся новые. В ходе наблюдений, проведённых в 1976—1979 годах, птицами было уничтожено 87 гнёзд без спутников. Только пять были восстановлены и впоследствии смогли дать потомство. В те же годы 12 гнёзд имели гнёзда-спутники, когда они были подвергнуты нападению птиц. Самки из восьми из них дали потомство, присоединившись к оставшемуся гнезду. Увеличение числа ос на гнезде не обеспечивает дополнительной защиты, как сокрытие гнезда и гнездо-спутник, которые снижают потери от нападения птиц[26].
Нападение муравья Crematogaster laeviuscula отличается от нападения птиц. Вместо того чтобы разрушить гнездо, муравьи роем облепляют его и удаляют из него расплод. Муравьи уничтожают весь расплод, но не могут убить взрослых особей ос[26].
Другие хищники нападают на взрослых особей, когда те кормятся или перемещаются к гнёздам-спутникам. Трудно определить количество взрослых особей, погибших вдали от гнезда, поскольку их трудно отследить. Однако было замечено, что полёт королевы к гнезду-спутнику не представляет опасности и не снижает её выживаемости[29]. Взрослых особей ос чаще всего убивают несколько видов хищных мух-ктырей (Diogmites angustipennis[англ.], Asilidae) и пауки-кругопряды (Araneidae)[26].
Идея групповой жизни обсуждалась многими эволюционистами с точки зрения её издержек и преимуществ. Было предсказано, что издержки больших групп заключаются в том, что они чаще страдают от паразитизма, но в то же время они выигрывают за счёт защиты от хищников. Эта теория не подтверждается в случае P. exclamans, поскольку паразитизм не всегда увеличивается с размером гнезда, а уровень хищничества не зависит от размера[30].

 Diogmites angustipennis
Diogmites angustipennis
Защита колонии

Лицевые узоры
Некоторые животные могут оценивать соперников по специфическим признакам, которые имеют отношение к их боевым способностям; однако некоторые животные вместо этого используют обычные признаки для определения конкурентных способностей соперников[31]. Обычные признаки — это сигналы, которые указывают на способности соперника, но не являются отражением физиологических или физических агонистических способностей животного[31]. P. exclamans использует рисунок лица в качестве обычного признака для определения агонистических способностей соперников. Более крупные осы имеют более пигментированных коричневый клипеус. Осы используют эти пигментированные клипеусы, чтобы определить, стоит ли бросать вызов сопернику[31]. Чем больше пигментированы клипеусы или чем крупнее P. exclamans, тем больше вероятность того, что P. exclamans захочет бросить вызов сопернику. Чем менее пигментированы клипеус или чем меньше P. exclamans, тем менее охотно она бросит вызов сопернику. Это помогает минимизировать «издержки конфликта во время конкуренции за доминирование среди гнездовых королев»[31].
Реакция тревоги
Polistes exclamans демонстрируют реакцию тревоги, характерную для многих высших эусоциальных перепончатокрылых (Hymenoptera)[32]. Хотя ранее указывалось, что соплеменники по гнезду могут тревожить остальных особей движениями рывками и жужжанием крыльев, P. exclamans способен оповещать остальных особей и привлекать атакующих ос химическими средствами, выпуская не видовой ядовитый сигнальный феромон[32]. Хотя химический сигнал, предупреждающий об атаке, был бы эволюционно адаптивным, P. exclamans подаёт ответный сигнал только после первоначального нападения хищника, поскольку яд выделяется только во время ужаления. Этот яд способен предупредить и скоординировать ответную реакцию из гнезда и привлечь близлежащих гетероспецифических или конспецифических самок из соседних колоний для нападения на хищника[32]. Это помогает удержать хищника от дальнейшего нападения и является особенно адаптивным для защиты гнезда. Хотя некоторые другие насекомые смогли приспособить выделение феромона к другим способам передачи сигнала тревоги, P. exclamans ещё не сделал этого, что указывает на то, что он все ещё находится в «примитивном состоянии в эволюции сложных систем коммуникации тревоги, наблюдаемых у высших социальных насекомых»[32]. В одном эксперименте феромоны были извлечены из желёз самок и нанесены на бумагу. Затем было обнаружено, что самки привлекались к ядовитой бумаге, а некоторые даже пытались её ужалить. При этом наблюдалась гетероспецифическая реакция, когда другие особи также реагировали на нее. Хотя было установлено, что одна оса не может выпустить достаточно феромонов, чтобы вызвать ответную реакцию, если многие особи настораживаются, они вызывают ответную реакцию у остальных членов гнезда. Однако этот запах недостаточно силён, чтобы достичь других гнёзд, поэтому соседние гнёзда должны быть встревожены физическими движениями и хлопаньем крыльев[32].
Реакция королевы
После вторжения королева может быть вынуждена покинуть своё гнездо. Если гнездо-спутник не было создано, королева может узурпировать другую королеву в другом гнезде. Не вкладывая никаких ценных ресурсов, королева может использовать ресурсы нового гнезда для воспроизводства нового выводка[33].
Примечания
- ↑ 1 2 3 Carpenter J. M. Distributional checklist of the species of the genus Polistes (Hymenoptera: Vespidae; Polistinae, Polistini) (англ.) // American Museum Novitates : Журнал. — 1996. — P. 1–39.
- ↑ 1 2 3 4 5 West M. J. Range extension and solitary nest founding in Polistes exclamans (англ.) // Psyche : Журнал. — 1968. — Vol. 75. — P. 118–123. — doi:10.1155/1968/49846.
- ↑ 1 2 3 4 Buck, Matthias Identification Atlas of Vespidae (англ.). University of Alberta. Дата обращения: 9 января 2013.
- ↑ 1 2 3 4 5 6 7 Strassmann J. E. Worker mortality and the evolution of castes in the social wasp, Polistes exclamans (англ.) // Insectes Sociaux[англ.] : Журнал. — 1985. — Vol. 32. — P. 275–285. — doi:10.1007/BF02224916.
- ↑ Viereck H. L. Notes and Descriptions of Hymenoptera from the Western United States (англ.) // Transactions of the American Entomological Society : Журнал. — 1906. — Vol. 32. — P. 173—246 (201—202).
- ↑ 1 2 Arevalo Elisabeth, Zhu Yong, Carpenter James, Strassmann Joan. The Phylogeny of the Social Wasp Subfamily Polistinae: Evidence from Microsatellite Flanking Sequences, Mitochondrial COIsequence, and Morphological Characters (англ.) // BMC Evolutionary Biology : Журнал. — 2004. — Vol. 4. — doi:10.1186/1471-2148-4-8. — PMC 385225.
- ↑ 1 2 3 4 5 Bequaert Joseph C. An Introductory Study of Polistes in the United States and Canada with Descriptions of Some New North and South American Forms (Hymenoptera: Vespidae) (англ.) // Journal of the New York Entomological Society : Журнал. — 1940. — Vol. 48. — P. 1–31.
- ↑ 1 2 Buck M., Marshall S. A., D. K. B. Cheung. Identification Atlas of the Vespidae (Hymenoptera, Aculeata) of the northeastern Nearctic region (англ.). — 2008. — P. 313—314. — 492 p. — (Canadian Journal of Arthropod Identification No.5). — doi:10.3752/cjai.2008.05.
- ↑ 1 2 3 4 Strassmann J. E. and M. C. Ferreira Orgren. Nest Architecture and Brood Development Times in the Paper Wasp, Polistes Exclamans (Hymenoptera: Vespidae) (англ.) // Psyche : Журнал. — 1983. — Vol. 90, no. 3. — P. 237—248. — doi:10.1155/1983/32347.
- ↑ Reed H.C. and S.B. Vinson. Nesting Ecology of Paper Wasps (Polistes) in a Texas Urban Area (Hymenoptera: Vespidae) (англ.) // Journal of the Kansas Entomological Society : Журнал. — 1979. — Vol. 52, no. 4. — P. 673—689.
- ↑ 1 2 3 4 5 Solis C. R., J. E. Strassmann. Presence of brood affects caste differentiation in the social wasp, Polistes exclamans Viereck (Hymenoptera: Vespidae) (англ.) // Functional Ecology : Журнал. — 1990. — Vol. 4, no. 4. — P. 531—541.
- ↑ 1 2 3 4 5 6 7 Strassman Joan E. Evolutionary implications of early male and satellite nest production in Polistes exclamans colony cycles (англ.) : Журнал. — 1981. — Vol. 8. — P. 55–64. — doi:10.1007/BF00302844.
- ↑ 1 2 3 4 Strassmann Joan E. Female-Biased Sex Ratios in Social Insects Lacking Morphological Castes (англ.) // Evolution : Журнал. — 1984. — Vol. 38. — P. 256–266.
- ↑ Eickwort K. R. Differential variation of males and females in Polistes exclamans (англ.) // Evolution : Журнал. — 1969. — Vol. 23. — P. 391–405. — doi:10.2307/2406695. — PMID 28562926. — .
- ↑ 1 2 Strassmann Joan E., Lee R. E. Jr., Rojas R. R., Baust J. G. Caste and sex differences in cold-hardiness in the social wasps, Polistes annularis and P. exclamans (англ.) // Insectes Sociaux[англ.] : Журнал. — 1984. — Vol. 31. — P. 291–301. — doi:10.1007/BF02223613.
- ↑ 1 2 3 4 Bohm M. K. Effects of environment and juvenile hormone on ovaries of the wasp, Polistes metricus (англ.) // Insect Physiology : Журнал. — 1972. — Vol. 18. — P. 1875–1883. — doi:10.1016/0022-1910(72)90158-8. — .
- ↑ 1 2 3 4 5 6 7 Strassmann Joan E., Meyer Dana C. Gerontocracy in the social wasp, Polistes exclamans (англ.) // Animal Behaviour : Журнал. — 1983. — Vol. 31. — P. 431–438. — doi:10.1016/S0003-3472(83)80063-3.
- ↑ 1 2 Herman R., Queller D., Strassmann Joan E. The role of queens in colonies of the swarm-founding wasp Parachartergus colobopterus (англ.) // Animal Behaviour : Журнал. — 2000. — Vol. 59. — P. 841–848. — doi:10.1006/anbe.1999.1385. — PMID 10792939.
- ↑ Strassmann, Joan What we have figured out about social insects? (англ.). The Queller/Strassmann Lab. Washington University in St. Louis (22 февраля 2012). Дата обращения: 21 сентября 2024.
- ↑ Lester L. J., R. K. Selander. Population Genetics of Haplodiploid Insects (англ.) // Genetics[англ.] : Журнал. — 1979. — Vol. 92. — P. 1329–1345. — doi:10.1093/genetics/92.4.1329. — PMID 17248955. — PMC 1214074.
- ↑ 1 2 3 Reed H. C., P. J. Landolt. Sex attraction in paper wasps, Polistes exclamans Viereck, in a wind tunnel (англ.) // Journal of Chemical Ecology : Журнал. — 1990. — Vol. 16. — P. 1277–1287. — doi:10.1007/BF01021026. — PMID 24263727.
- ↑ 1 2 Allen J. L., K. Schulze-Kellman, and G. J. Gamboa. Clumping Patterns During Overwintering in the Paper Wasp, Polistes exclamans: effects of Relatedness (англ.) // Journal of the Kansas Entomological Society : Журнал. — 1982. — Vol. 55, no. 1. — P. 97—100.
- ↑ 1 2 Gonzalez J. M., S. B. Vinson. Does Polistes exclamans Vierek (Hymenoptera: Vespidae) Hibernate Inside Muddauber Nests (англ.) // Southwestern Entomologist : Журнал. — 2007. — Vol. 32, no. 1. — P. 67—71.
- ↑ Judd T. M. Division of labour in colony defence against vertebrate predators by the social wasp Polistes fuscatus (англ.) // Animal Behaviour : Журнал. — 2000. — Vol. 60. — P. 55–61. — doi:10.1006/anbe.2000.1449. — PMID 10924203.
- ↑ Krebs J. R., Nicholas B. Davies. Living in Groups // An Introduction to Behavioral Ecology. 4th ed. Vol. 1. — Oxford: Blackwell Scientific Publications, 1989. — С. 147—178.
- ↑ 1 2 3 4 5 Strassmann Joan E. Parasitoids, predators, and group size in the paper wasp, Polistes exclamans (англ.) // Ecology : Журнал. — 1981. — Vol. 62. — P. 1225–1233. — doi:10.2307/1937287. — . — .
- ↑ 1 2 Lutz G., Strassmann J., Hughes C. Nest Defense by the social wasps Polistes exclamans and P instabilis (англ.) // Entomological News : Журнал. — 1984. — Vol. 95. — P. 47–50. — doi:10.1007/BF00302844.
- ↑ Strassmann J., Thomas R. An analysis of the interrelationships among nest variables in Polistes exclamans (англ.) // Journal of the Kansas Entomological Society : Журнал. — 1980. — Vol. 53. — P. 770–780. — .
- ↑ Seppa P., Queller D., Strassmann J. Reproduction in foundress associations of the social wasp, Polistes carolina: conventions, competition, and skew. (англ.) // Behavioral Ecology[англ.] : Журнал. — 1892. — Vol. 13. — P. 531–542. — doi:10.1093/beheco/13.4.531.
- ↑ Alexander R., Borgia G. Group Selection, Altruism, and the Levels of Organization of Life (англ.) // Annual Review of Ecology and Systematics : Журнал. — 1978. — Vol. 9. — P. 449–474. — doi:10.1146/annurev.es.09.110178.002313.
- ↑ 1 2 3 4 Tibbetts E. A., M. J. Sheehan. Facial Patterns are a Conventional Signal of Agonistic Ability in Polistes exclamans Paper Wasps (англ.) // Ethology : Журнал. — 2011. — Vol. 117, no. 12. — P. 1138—1146. — doi:10.1111/j.1439-0310.2011.01967.
- ↑ 1 2 3 4 5 Post D., Downing H., Jeanne R. Alarm response to venom by social wasps Polistes exclamans and P. fuscatus (англ.) // Journal of Chemical Ecology : Журнал. — 1984. — Vol. 10. — P. 1425–1433. — doi:10.1007/BF00990313. — PMID 24318343.
- ↑ Cervo R. Polistes wasps and their social parasites: an overview (англ.) // Ann. Zool. Fennici : Журнал. — Finnish Zoological and Botanical Publishing Board, 2006. — Vol. 43. — P. 531–549. — ISSN 0003-455X.
Литература
- Carpenter J. M. Distributional checklist of the species of the genus Polistes (Hymenoptera: Vespidae; Polistinae, Polistini) (англ.) // American Museum Novitates : Журнал. — 1996. — P. 1–39.
- Richards O. W. The social wasps of the Americas excluding the Vespinae. — London: British Museum (Natural History), 1978. — 580 p.
| В библиографических каталогах |
|---|