SMC3


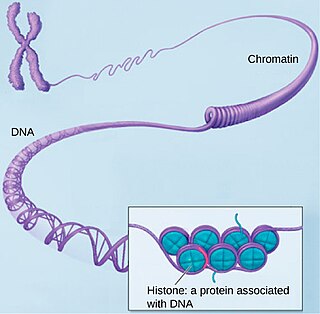
SMC3 или белок структурной поддержки хромосом номер три (англ. Structural maintenance of chromosomes protein 3) — белок, который у человека кодируется геном SMC4[1].
Длина полипептидной цепи белка составляет 1217 аминокислотных остатков, а молекулярная масса — 141542 Да[2].
Структура и взаимодействия


Доменная организация SMC-белков эволюционно консервативна и состоит из N-концевого мотива Walker A, катушки (двуспиральная область), "петли", катушки и С-концевого мотива Walker B. Белок складывается сам на себя, образуя стержнеобразную молекулу с гетеродимеризационным "петлевым" доменом на одном конце и АТФазной "головкой" ABC-типа на другом. Эти глобулярные домены разделены антипараллельной спиралью длиной ~ 50 нм. SMC3 и SMC1 связываются через свои шарнирные домены, образуя V-образные гетеродимеры. N-концевой домен RAD21 связывается со спиралью SMC3 чуть выше головного домена, а C-концевой домен RAD21 связывается с головным доменом SMC1. В результате такого связывания тримера SMC3-SMC1-RAD21 образуется замкнутое кольцо, внутри которого может быть захвачена ДНК.
Когда происходит репликация ДНК и устанавливается сцепление сестринских хроматид, SMC3 ацетилируется по паре высококонсервативных лизинов под действием ESCO1 и ESCO2. У почкующихся дрожжей этой модификации достаточно для стабилизации когезина на ДНК до митоза, но у животных также требуется связывание сорорина.
Во время мейоза SMC3 образует когезиновые комплексы с SMC1ß, STAG3 и REC8, которые создают когезию между гомологичными хромосомами и сестринскими хроматидами[3].
Функции
Этот ген принадлежит к SMC3 подсемейству SMC-белков. Белок, кодируемый этим геном, может присутствовать в определённых типах клеток как внутриклеточный, ядерный или даже секретируемый белок. Ядерная форма, известная как белок структурной поддержки хромосом 3, является компонентом когезина, мультибелкового комплекса, который скрепляет сестринские хроматиды в процессе митоза, обеспечивая надлежащую сегрегацию хромосом. По окончании синтеза этот белок может подвергаться посттрансляционной модификации — добавлению цепи хондроитин сульфата. В результате образуется секретируемый протеогликан бамакан, который в больших количествах содержится в базальной мембране[1].
Синдром Корнелии де Ланге
Синдром Корнелии де Ланге (CdLS) — редкое генетическое заболевание, которое проявляется различными клиническими аномалиями, включая дисморфические черты, тяжёлую задержку роста, глобальную задержку развития и умственную отсталость. SMC3 —— один из пяти генов, которые были связаны с CdLS[4]. В одном из сообщений о случае была обнаружена новая дупликация гена SMC3 у ребёнка с задержкой роста, гипотонией и дисморфическими чертами лица при синдроме Корнелии де Ланге[4]. Такая же дупликация была обнаружена и у матери, у которой дисморфические черты лица были более мягкими.
Модельные организмы
| Характеристика | Фенотип |
|---|---|
| Жизнеспособность гомозигот | ненормальный |
| Летальность для гомозигот | ненормальный |
| Фертильность гомозигот | ненормальный |
| Вес тела | ненормальный[5] |
| Беспокойность | Норма |
| Неврология | Норма |
| Сила хватки | Норма |
| Горячая тарелочка | Норма |
| Тератология | ненормальный |
| Непрямая калориметрия | Норма |
| Глюкозотолерантный тест | Норма |
Слуховой ответ ствола мозга | Норма |
| DEXA | ненормальный |
| Рентгенография | Норма |
| Температура тела | Норма |
| Морфология глаза | Норма |
| Клиническая химия | Норма |
| Гематология | Норма |
| Периферические лимфоциты | Норма |
| Масса сердца | Норма |
| Гистопатологии глаза | Норма |
| Инфицирование Salmonella | Норма[6] |
| Инфицирование Citrobacter | Норма[7] |
| Информация взята из[8][9] |
В исследовании функции гена SMC3 были использованы нокаутные мыши линии Smc3tm1a(EUCOMM)Wtsi[10][11]. Линия была создана в рамках Международного консорциума нокаут мыши программа — с высокой пропускной способностью анализа мутантов животных, представляющих модели заболеваний, которые интересуют учёных[12][13][14].
Особей мужского и женского пола прошли стандартизированный фенотипические тесты, чтобы определить последствия делеции гена[8][15]. Двадцать два испытания проводили на мутантных мышей и было обнаружено шесть существенных отклонений[8]. Гомозиготы по этой мутации погибали ещё нас ранних стадиях эмбриогенеза. Все тесты проводились на гетерозиготных взрослых мышах. У самок наблюдалась повышенная смертность в потомстве до перехода к самостоятельному питанию, а также снижение массы тела. Самцы гетерозигот имели укороченную, вздернутую морду[8][15].
См. также
Примечания
- ↑ 1 2 Entrez Gene: SMC3 structural maintenance of chromosomes 3. Архивировано 7 марта 2010 года.
- ↑ UniProt, Q9UQE7 (англ.). Дата обращения: 19 декабря 2023. Архивировано 19 декабря 2023 года.
- ↑ Garcia-Cruz R, Brieño MA, Roig I, Grossmann M, Velilla E, Pujol A, Cabero L, Pessarrodona A, Barbero JL, Garcia Caldés M (2010). "Dynamics of cohesin proteins REC8, STAG3, SMC1 beta and SMC3 are consistent with a role in sister chromatid cohesion during meiosis in human oocytes". Hum. Reprod. 25 (9): 2316—27. doi:10.1093/humrep/deq180. PMID 20634189.
- ↑ 1 2 Infante E, Alkorta-Aranburu G, El-Gharbawy A (2017). "Rare form of autosomal dominant familial Cornelia de Lange syndrome due to a novel duplication in SMC3". Clinical Case Reports. 5 (8): 1277—1283. doi:10.1002/ccr3.1010. PMC 5538066. PMID 28781842.
- ↑ Body weight data for Smc3. Wellcome Trust Sanger Institute. Дата обращения: 17 ноября 2015. Архивировано 17 октября 2012 года.
- ↑ Salmonella infection data for Smc3. Wellcome Trust Sanger Institute.
- ↑ Citrobacter infection data for Smc3. Wellcome Trust Sanger Institute. Дата обращения: 17 ноября 2015. Архивировано 17 октября 2012 года.
- ↑ 1 2 3 4 Gerdin A.K. The Sanger Mouse Genetics Programme: High throughput characterisation of knockout mice (англ.) // Acta Ophthalmologica[англ.] : journal. — 2010. — Vol. 88. — P. 925—927. — doi:10.1111/j.1755-3768.2010.4142.x.
- ↑ Mouse Resources Portal Архивная копия от 24 декабря 2011 на Wayback Machine, Wellcome Trust Sanger Institute.
- ↑ International Knockout Mouse Consortium. Дата обращения: 17 ноября 2015. Архивировано из оригинала 3 апреля 2012 года.
- ↑ Mouse Genome Informatics. Дата обращения: 17 ноября 2015. Архивировано 18 ноября 2015 года.
- ↑ Skarnes W.C., Rosen B., West A.P., Koutsourakis M., Bushell W., Iyer V., Mujica A.O., Thomas M., Harrow J., Cox T., Jackson D., Severin J., Biggs P., Fu J., Nefedov M., de Jong P.J., Stewart A.F., Bradley A. A conditional knockout resource for the genome-wide study of mouse gene function (англ.) // Nature : journal. — 2011. — Vol. 474, no. 7351. — P. 337—342. — doi:10.1038/nature10163. — PMID 21677750. — PMC 3572410.
- ↑ Dolgin E. Mouse library set to be knockout (англ.) // Nature. — 2011. — Vol. 474, no. 7351. — P. 262—263. — doi:10.1038/474262a. — PMID 21677718.
- ↑ Collins F.S., Rossant J., Wurst W. A mouse for all reasons (англ.) // Cell. — Cell Press, 2007. — Vol. 128, no. 1. — P. 9—13. — doi:10.1016/j.cell.2006.12.018. — PMID 17218247.
- ↑ 1 2 van der Weyden L., White J.K., Adams D.J., Logan D.W. The mouse genetics toolkit: revealing function and mechanism (англ.) // Genome Biology[англ.] : journal. — 2011. — Vol. 12, no. 6. — P. 224. — doi:10.1186/gb-2011-12-6-224. — PMID 21722353. — PMC 3218837.