
Рибонуклеи́новая кислота́ (РНК) — одна из трёх основных макромолекул, которые содержатся в клетках всех живых организмов и играют важную роль в кодировании, прочтении, регуляции и экспрессии генов.

РНК-интерференция — процесс подавления экспрессии гена на стадии транскрипции, трансляции, деаденилирования или деградации мРНК при помощи малых молекул РНК.

РНК-полимераза — фермент, осуществляющий синтез молекул РНК. В узком смысле, РНК-полимеразой обычно называют ДНК-зависимые РНК-полимеразы, осуществляющие синтез молекул РНК на матрице ДНК, то есть осуществляющие транскрипцию. Ферменты класса РНК-полимераз очень важны для функционирования клетки, поэтому они имеются во всех организмах и во многих вирусах. Химически РНК-полимеразы являются нуклеотидил-трансферазами, полимеризующими рибонуклеотиды на 3'-конце цепи РНК.

Ми́кроРНК — малые некодирующие молекулы РНК длиной 18—25 нуклеотидов, обнаруженные у растений, животных и некоторых вирусов, принимающие участие в транскрипционной и посттранскрипционной регуляции экспрессии генов путём РНК-интерференции. Помимо внутриклеточной обнаружена внеклеточная (циркулирующая) микроРНК.
Виро́иды — инфекционные агенты, состоящие только из кольцевой РНК. Они вызывают различные болезни растений, в том числе веретеновидность клубней картофеля, экзокортис цитрусовых и карликовость хризантемы. По оценкам учёных, более трети вирусных заболеваний растений вызываются вироидами.

Некодирующие РНК — молекулы РНК, которые не транслируются в белки. Ранее использовавшийся синоним — «малые РНК» — вышел из употребления, так как некоторые некодирующие РНК могут быть очень большими, например, Xist.
Ма́лые интерфери́рующие РНК или короткие интерферирующие РНК — это класс двухцепочечных РНК, длиной 20-25 нуклеотидов. Взаимодействие малых интерферирующих РНК с матричной РНК (мРНК) целевого гена приводит к деградации последней, предотвращая трансляцию мРНК на рибосомах в кодируемый ею белок. В конечном итоге результат действия малых интерферирующих РНК идентичен тому, как если бы просто снижалась экспрессия гена.
piРНК — наиболее крупный класс малых некодирующих РНК, экспрессируемых в клетках животных; они обнаружены в комплексах с белками семейства Piwi, за что и получили своё название. piРНК обычно длиннее микроРНК и малых интерферирующих РНК и имеют длину 26—32 нуклеотида, кроме того, в отличие от микроРНК, они не так консервативны. Белки Piwi относятся к большой группе белков Argonaute и экспрессируются почти исключительно в клетках зародышевой линии; они необходимы для поддержания стволовых клеток зародышевой линии, сперматогенеза и репрессии мобильных элементов. Комплексы Piwi с piРНК не только задействованы в сайленсинге ретротранспозонов и других генетических элементов на пост-трансляционном уровне, но имеют и некоторые другие, в значительной мере ещё неописанные эффекты, например, эпигенетические.
Малые ядрышковые РНК — класс малых РНК, участвующих в химических модификациях рибосомных РНК, а также тРНК и малых ядерных РНК. По классификации MeSH малые ядрышковые РНК считаются подгруппой малых ядерных РНК. мякРНК обычно относят к «гидовым» РНК, однако их нельзя путать с гидовыми РНК, направляющими редактирование РНК у трипаносом.

Антисмысловы́е РНК — одноцепочечные РНК, которые комплементарны мРНК, транскрибируемой в клетке, или гену-мишени. Механизмы действия антисмысловых РНК весьма разнообразны, они могут как подавлять, так и активировать экспрессию гена-мишени. Природные антисмысловые РНК есть и у прокариот, и у эукариот; они относятся к длинным некодирующим РНК как РНК длиной более 200 нуклеотидов. Синтетические антисмысловые РНК нашли широкое применение у исследователей в качестве инструмента для нокдауна генов. Антисмысловые РНК также находят медицинское применение.
Активация РНК — процесс активации экспрессии генов короткими двуцепочечными РНК. Известно, что короткие двуцепочечные РНК (дцРНК) могут снижать экспрессию генов по эволюционно древнему механизму, называемому РНК-интерференцией. Такие короткие дцРНК называют малыми интерферирующими РНК. Явление РНК-интерференции может происходить как на уровне транскрипции, так и на посттранскрипционном уровне. Результаты исследований, опубликованные в 2006-2008 годах указывают на то, что наряду со снижением, короткие дцРНК могут также активировать экспрессию генов.

Шпи́лька — в молекулярной биологии элемент вторичной структуры РНК, а также одноцепочечной ДНК. Шпилька образуется в том случае, когда две последовательности одной и той же цепи комплементарны друг другу и соединяются друг с другом, перегибаясь одна к другой и образуя на конце неспаренный участок — петлю. Такие комплементарные последовательности нередко представляют собой палиндромные последовательности.
Y-РНК — малые некодирующие РНК, входят в состав рибонуклеопротеинов, содержащих белки Ro60 и La, которые являются мишенью аутоантител у пациентов, страдающих системной красной волчанкой и синдромом Шегрена. Они также необходимы для репликации ДНК, так как взаимодействуют с хроматином и инициаторными белками.
Транс-активи́рующие ма́лые интерфери́рующие РНК, tasiРНК, TAS РНК — группа малых некодирующих РНК наземных растений, подавляющие экспрессию генов путём пост-трансляционного сайленсинга. TasiРНК транскрибируются в геноме в форме двуцепочечных полиаденилированных РНК, которые в дальнейшем процессируются и превращаются во фрагменты РНК длиной 21 нуклеотид. Эти фрагменты включаются в РНК-индуцируемый комплекс выключения гена (RISC). tasiРНК часто относят к малым интерферирующим РНК (siРНК) ввиду того, что обе этих группы малых РНК транскрибируются в форме двуцепочечных РНК и подвергаются схожему процессингу. Впрочем, tasiРНК отличаются от других siРНК тем, что они связывают свои последовательности-мишени с меньшей специфичностью. В этом их механизм более схож с механизмом действия микроРНК, так как они не нуждаются в полной комплементарности последовательностей со своей мишенью, чтобы направлять её распад.

Внутренняя терминация, также ρ-независимая терминация, ро-независимая терминация , — механизм остановки транскрипции гена у прокариот. Суть этого механизма такова: мРНК содержит гуанин-цитозин-обогащённую последовательность, которая может образовывать структуры типа шпилька в 7—20 пар оснований в длину. Гуанин и цитозин образуют друг с другом три водородные связи и поэтому связаны довольно прочно. Сразу после шпильки располагается участок, обогащённый урацилом. Связи между урацилом и аденином очень слабы. Белок, связанный с РНК-полимеразой (nusA), настолько прочно связывается со шпилькой, что это вызывает временную остановку полимеразы и прекращение транскрипции. В этот момент полимераза располагается на полиурациловом участке последовательности. Слабые аденин-урациловые связи снижают энергию дестабилизации дуплекса РНК-ДНК, что позволяет им ослабить напряжение полинуклеотидной цепи и диссоциировать от РНК-полимеразы.

РНК-полимераза II — фермент эукариот, который катализирует транскрипцию ДНК, синтезирует предшественников мРНК и большинство мяРНК и микроРНК. Эта полимераза представляет собой комплекс массой 550 кДа, состоящий из 12 субъединиц. РНК-полимераза II является наиболее изученным типом РНК-полимеразы. Ей необходим широкий спектр транскрипционных факторов для того, чтобы связываться с генами выше промоторов и начинать транскрипцию.
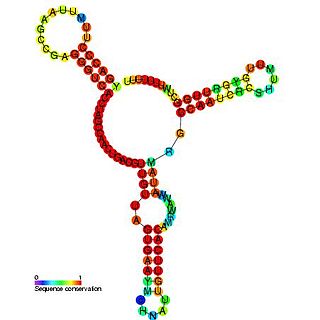
Ма́лые РНК бакте́рий — небольшие некодирующие РНК длиной 50—250 нуклеотидов, содержащиеся в клетках бактерий. Как правило, малые РНК бактерий имеют сложную структуру и содержат несколько шпилек. Многочисленные малые РНК были определены в клетках кишечной палочки, модельном патогене Salmonella, азотфиксирующей альфа-протеобактерии Sinorhizobium meliloti, морских цианобактериях, возбудителе туляремии Francisella tularensis, патогене растений Xanthomonas oryzae pathovar oryzae и других бактериях. Для поиска малых РНК в геноме бактерий использовали компьютерный анализ и различные лабораторные методы.
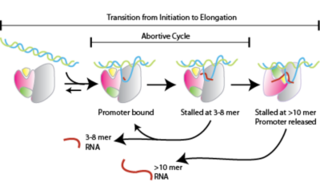
Абортивная инициация, также известная как абортивная транскрипция, ранний процесс генетической транскрипции, в котором РНК-полимераза связывается с промотором ДНК и входит в циклы синтеза коротких транскриптов мРНК, которые выделяются прежде, чем транскрипционный комплекс покинет промотор. Этот процесс происходит как у эукариот так и у прокариот. Абортивная инициация, как правило, изучается в РНК-полимеразах Т3 и Т7 в бактериофагах и кишечной палочке.
RITS — форма РНК-интерференции, при которой короткие молекулы РНК, такие как малые интерферирующие РНК (siРНК), подавляют транскрипцию гена-мишени. Это часто сопровождается посттрансляционными модификациями хвостов гистонов, а именно метилированием лизина 9 гистона H3 (H3K9me), которое приводят к образованию гетерохроматина в локусе-мишени. Таким образом, RITS участвует в образовании гетерохроматина de novo. Белковый комплекс, который связывается с siРНК и взаимодействует с метилированным остатком лизина 9 гистона Н3, называется комплексом RITS. RITS был открыт у делящихся дрожжей Schizosaccharomyces pombe, и было показано, что он принимает участие в инициации образования гетерохроматина и его поддержании в локусе типа спаривания и в образовании центромеры. В состав комплекса RITS S. pombe входят три белка: белок группы argonaute, содержащий piwi-домен и похожий на РНКазу Н, белок Chp1, содержащий хромодомен, и белок Tas3, взаимодействующий с белками argonaute и с Chp1. Для образования гетерохроматина необходимы, как минимум, белок argonaute и РНК-зависимая РНК-полимераза. Утрата генов, кодирующих эти белки, у S. pombe, приводит к нарушениям в структуре гетерохроматина и функционировании центромер, так как комплекс RITS содержит siРНК, считанную с центромерных повторов. Аномальное функционирование центромер, в свою очередь, приводит к нарушению сегрегации хромосом в митозе, а именно — к появлению «отстающих» хромосом на стадии анафазы.
В клетках эукариот РНК-полимераза III транскрибирует ДНК для синтеза рибосомной 5S рРНК, тРНК, 7SL РНК и других малых РНК.