
Рибонуклеи́новая кислота́ (РНК) — одна из трёх основных макромолекул, которые содержатся в клетках всех живых организмов и играют важную роль в кодировании, прочтении, регуляции и экспрессии генов.

РНК-интерференция — процесс подавления экспрессии гена на стадии транскрипции, трансляции, деаденилирования или деградации мРНК при помощи малых молекул РНК.
Термина́тор — нуклеотидная последовательность ДНК, на которой завершается транскрипция гена или оперона. Как правило, последовательность терминатора такова, что комплементарная ей последовательность в мРНК вызывает выход новосинтезированного транскрипта из транскрипционного комплекса. Эта последовательность в мРНК может сама по себе вызывать терминацию за счёт собственной вторичной структуры, а может привлекать особые белки — факторы терминации. После высвобождения РНК-полимераза и транскрипционные факторы приступают к транскрипции другого гена.

РНК-полимераза — фермент, осуществляющий синтез молекул РНК. В узком смысле, РНК-полимеразой обычно называют ДНК-зависимые РНК-полимеразы, осуществляющие синтез молекул РНК на матрице ДНК, то есть осуществляющие транскрипцию. Ферменты класса РНК-полимераз очень важны для функционирования клетки, поэтому они имеются во всех организмах и во многих вирусах. Химически РНК-полимеразы являются нуклеотидил-трансферазами, полимеризующими рибонуклеотиды на 3'-конце цепи РНК.
Малые ядерные РНК — класс РНК, которые встречаются в ядре эукариотических клеток. Они транскрибируются РНК-полимеразой II или РНК-полимеразой III и участвуют в важных процессах, таких как сплайсинг, регуляции факторов транскрипции или РНК-полимеразы и поддержании целостности теломер.

Ми́кроРНК — малые некодирующие молекулы РНК длиной 18—25 нуклеотидов, обнаруженные у растений, животных и некоторых вирусов, принимающие участие в транскрипционной и посттранскрипционной регуляции экспрессии генов путём РНК-интерференции. Помимо внутриклеточной обнаружена внеклеточная (циркулирующая) микроРНК.
Белки группы polycomb — это семейство белков, которые способны ремоделировать хроматин. Эти белки-регуляторы были впервые описаны у дрозофил, где они подавляют гомеозисные гены, контролирующие индивидуальные отличия сегментов развивающегося эмбриона.
Полиаденили́рование — это процесс присоединения большого количества остатков аденозинмонофосфата к 3'-концу первичной мРНК (пре-мРНК). Иными словами, поли(А)-хвост — это фрагмент молекулы мРНК, азотистые основания которого представлены только аденином. У эукариот полиаденилирование является частью процессинга мРНК — процесса созревания первичного транскрипта в зрелую мРНК, готовую для трансляции. Процессинг, в свою очередь, является одним из этапов экспрессии генов.

Кэп, 5'-кэп, или кэп-структура — структура на 5'-конце матричных РНК (мРНК) и некоторых других РНК эукариот. Кэп состоит из одного или нескольких модифицированных нуклеотидов и характерен только для транскриптов, синтезируемых РНК-полимеразой II. Наличие кэпа — один из признаков, отличающих эукариотические мРНК от прокариотических, которые несут трифосфат на 5'-конце. Это и другие отличия обуславливают существенно более высокую стабильность, особый механизм инициации трансляции и другие особенности жизненного цикла эукариотической мРНК.

Экзосомы — микроскопические внеклеточные везикулы (пузырьки) диаметром 30—100 нанометров, выделяемые в межклеточное пространство клетками различных тканей и органов. Полость экзосом имеет цитоплазматическое происхождение и содержит белки, РНК и липиды, мембрана экзосом образуется в результате впячивания внутрь эндосомальной мембраны.

Экзосо́ма (ко́мплекс), или PM/Scl-ко́мплекс — мультибелковый комплекс, способный к разрушению различных типов молекул РНК. Экзосомы имеются у эукариот и архей, в то время как у бактерий сходные функции выполняет более просто устроенный комплекс — деградосома.

Обрабатывающий сплайсинг пре-мРНК фактор 8 — белок, который у человека кодируется геном PRPF8.
Транс-активи́рующие ма́лые интерфери́рующие РНК, tasiРНК, TAS РНК — группа малых некодирующих РНК наземных растений, подавляющие экспрессию генов путём пост-трансляционного сайленсинга. TasiРНК транскрибируются в геноме в форме двуцепочечных полиаденилированных РНК, которые в дальнейшем процессируются и превращаются во фрагменты РНК длиной 21 нуклеотид. Эти фрагменты включаются в РНК-индуцируемый комплекс выключения гена (RISC). tasiРНК часто относят к малым интерферирующим РНК (siРНК) ввиду того, что обе этих группы малых РНК транскрибируются в форме двуцепочечных РНК и подвергаются схожему процессингу. Впрочем, tasiРНК отличаются от других siРНК тем, что они связывают свои последовательности-мишени с меньшей специфичностью. В этом их механизм более схож с механизмом действия микроРНК, так как они не нуждаются в полной комплементарности последовательностей со своей мишенью, чтобы направлять её распад.

РНК-полимераза II — фермент эукариот, который катализирует транскрипцию ДНК, синтезирует предшественников мРНК и большинство мяРНК и микроРНК. Эта полимераза представляет собой комплекс массой 550 кДа, состоящий из 12 субъединиц. РНК-полимераза II является наиболее изученным типом РНК-полимеразы. Ей необходим широкий спектр транскрипционных факторов для того, чтобы связываться с генами выше промоторов и начинать транскрипцию.
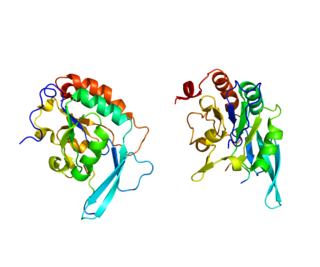
Субъединица RPB1 ДНК-управляемой РНК-полимеразы II , также известная как RPB1 , — фермент, кодируемый у человека геном POLR2A .

Субъединица RPB7 ДНК-управляемой РНК-полимеразы II — фермент, кодируемый у человека геном POLR2G .

Субъединица RPABC5 ДНК-управляемых РНК-полимераз I, II и III — белок, кодируемый у человека геном POLR2L .
RITS — форма РНК-интерференции, при которой короткие молекулы РНК, такие как малые интерферирующие РНК (siРНК), подавляют транскрипцию гена-мишени. Это часто сопровождается посттрансляционными модификациями хвостов гистонов, а именно метилированием лизина 9 гистона H3 (H3K9me), которое приводят к образованию гетерохроматина в локусе-мишени. Таким образом, RITS участвует в образовании гетерохроматина de novo. Белковый комплекс, который связывается с siРНК и взаимодействует с метилированным остатком лизина 9 гистона Н3, называется комплексом RITS. RITS был открыт у делящихся дрожжей Schizosaccharomyces pombe, и было показано, что он принимает участие в инициации образования гетерохроматина и его поддержании в локусе типа спаривания и в образовании центромеры. В состав комплекса RITS S. pombe входят три белка: белок группы argonaute, содержащий piwi-домен и похожий на РНКазу Н, белок Chp1, содержащий хромодомен, и белок Tas3, взаимодействующий с белками argonaute и с Chp1. Для образования гетерохроматина необходимы, как минимум, белок argonaute и РНК-зависимая РНК-полимераза. Утрата генов, кодирующих эти белки, у S. pombe, приводит к нарушениям в структуре гетерохроматина и функционировании центромер, так как комплекс RITS содержит siРНК, считанную с центромерных повторов. Аномальное функционирование центромер, в свою очередь, приводит к нарушению сегрегации хромосом в митозе, а именно — к появлению «отстающих» хромосом на стадии анафазы.
Контро́ль ка́чества мРНК — совокупность молекулярных механизмов, обеспечивающих отбраковывание дефектных мРНК и не допускающих их трансляцию. Механизмы контроля качества мРНК действуют на разных этапах биогенеза мРНК. Как правило, они приводят к тому, что дефектные мРНК оказываются помеченными специфическим образом и благодаря этому распознаются ферментами-нуклеазами, разрушающими их.

Nonstop-деграда́ция — механизм контроля качества мРНК, направленный на выявление мРНК, лишённых стоп-кодона, и предотвращение их трансляции. В ходе nonstop-деградации рибосома, которая успела значительно продвинуться в сторону 3'-конца мРНК, диссоциирует, и мРНК направляется в экзосомный комплекс или к РНКазе R для дальнейшего разрушения.
