Tataouinea
| † Tataouinea | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Клада: Клада: Подотряд: Клада: Клада: Инфраотряд: Надсемейство: Семейство: Подсемейство: Род: † Tataouinea | |||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Tataouinea Fanti et al., 2013 | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
| Геохронология Альбский век 113,0—100,5 млн лет
◄ Наше время◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание◄ Массовоепермское вымирание ◄ Девонское вымирание◄ Ордовикско-силурийское вымирание◄ Кембрийский взрыв | |||||||||||||||||||||||||||||||||||||||||
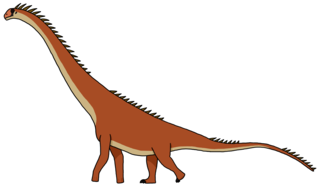
Tataouinea (лат.) — род зауроподовых динозавров семейства Rebbachisauridae, боковой ветви Diplodocoidea. Ископаемые остатки известны из нижнемеловых отложений Туниса (область Татавин). Род Tataouinea содержит один вид — Tataouinea hannibalis[1].
Подобно другим членам Rebbachisauridae, в числе которых хорошо изученный Nigersaurus, это, вероятно, был относительно некрупный четвероногий растительноядный динозавр с короткой шеей, длинным кнутовидным хвостом и лёгким телосложением. Обладая необычной респираторной системой, представители рода Tataouinea имели воздушные мешки, более характерные для птиц, чем для «нептичьих» динозавров[2].
Открытие и изучение

Осенью 2011 года на гранты университета Болоньи команда палеонтологов, в состав которой входил Фредерик Фанти, проводила исследования на юге Туниса в местности Джебель-эль-Мра (фр. Jebel El Mra). В ходе раскопок палеонтолог по имени Альдо Баккетта обнаружил ископаемые остатки крупного динозавра в подразделении Ум-эд-Диаб[англ.] (фр. Oum ed Diab) формации Айн-эль-Гуттер (фр. Aïn el Guetter)[1].
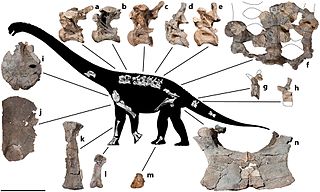
Обнаруженный командой Фанти типовой экземпляр (каталогизированный как ONM DT 1-36) состоял из фрагментов костей таза, нескольких хвостовых и крестцовых позвонков[1][3].
Первое описание вида было опубликовано в июльском номере журнала Nature Communications за 2013 год, авторы публикации — Фредерико Фанти, Андреа Кау и Микела Контесси из университета Болоньи; а также тунисский ученый Мохсен Хассине. Животное было описано под названием Tataouinea hannibalis, где родовое имя является отсылкой к месту находки вблизи города Татавин, а видовое название — к карфагенскому полководцу Ганнибалу (III—II века до нашей эры), участнику Второй Пунической войны[4].
В 2015 году в онлайн-издании PLOS One вышла статья, посвященная более детальному описанию каждой известной кости Tataouinea hannibalis (авторы публикации — Марко Аудиторе из палеонтологического музея Читтадино, Луиджи Кантелли из университета Болоньи, а также ранее упомянутые Фанти, Кау и Хассине). Статья содержала новую информацию об эволюции, палеобиогеографии и используемую в данный момент версию систематики, включающую кладу Khebbashia[англ.][5].
Описание


В связи с тем, что единственный экземпляр Tataouinea довольно плохо сохранился, исследователи, реконструируя прижизненный облик животного, опирались на описания видов, родственных Tataouinea hannibalis и Nigersaurus taqueti[3].
По оценкам Грегори Пола[англ.], Tataouinea была самым крупным животным семейства Rebbachisauridae после Limaysaurus tessonei[англ.][6]. Однако по меркам всего инфраотряда зауропод Tataouinea была, как и другие члены своего семейства, некрупным животным: при 14-метровой длине взрослая особь весила восемь тонн[7].
Tataouinea, как и все представители Rebbachisauridae, обладала, вероятно, короткой шеей (около 13—16 позвонков, как у Nigersaurus и Limaysaurus[8]), маленькой головой с широкими челюстями и длинным хвостом[6]. Передние конечности были тоньше и на треть короче задних[8].
Череп Tataouinea (как и у Nigersaurus) был, вероятно, пневматизирован посредством множества отверстий. Рыло было заметно шире затылка, челюсть динозавра, имевшая зубы только в передней части, напоминала пылесос[8].
Исследователи отмечают исключительно облегчённые кости посткраниального скелета, характерные также для Nigersaurus, так что тазовый пояс Tataouinea (как, вероятно, и плечевой) имел очень тонкие кости[8], их стенки имели в толщину всего один сантиметр. Исследователи отмечают наличие внутренних камер (англ. internal chambers) c септами в подвздошных костях и с нервно-сосудистыми отверстиями в седалищных костях[9]. На момент открытия Tataouinea подобных конструкций в седалищных костях других динозавров обнаружено не было[1].
Большая часть позвонков также имела внутри полости, заполненные губчатой тканью и воздухом, дуги позвонков были очень тонкими (не более двух миллиметров толщиной). У Tataouinea остистые отростки крестцовых позвонков были спаяны и создавали «парус» (горб) из мягких тканей, как у дикреозавра[7]. Общая длина крестца составляет 80 см[10]. Хвостовые позвонки были более цельными и не содержали столь крупных полостей. У позвонков, формирующих основание хвоста, имелись заметные эллиптические впадины. Крупные невральные отростки имели форму перевернутой буквы «Y»[11].
Необычно пневматизированные кости Tataouinea натолкнули исследователей на мысли о наличии у животного респираторной системы, схожей с современными птицами. Анатомические особенности скелета косвенно указывали на наличие внутри грудных и брюшных полостей воздушных мешков, стабильно обеспечивавших гигантское животное воздухом[2]. Так как у экземпляра ONM DT 1-36 крупные пневматические отверстия присутствуют и в костях таза, удаленных от прямого контакта с осевым скелетом, предположение о наличии «вентиляционной системы» внутри тазового пояса даже при наиболее консервативном подходе кажется правдоподобным[12].
Чаще всего воздушные мешки встречаются у птиц и некоторых птицеподобных динозавров, в частности, сообщения о таких дыхательных приспособлениях встречались в описании теропода Aerosteon[12]. До открытия Tataouinea о наличии подобных дыхательных систем сообщалось и у других видов зауроподовых динозавров[13][8].
Систематика
В первом описании от 2013 года Фанти и коллеги поместили Tataouinea в подсемейство Nigersaurinae[англ.] семейства Rebbahisauridae, боковой ветви надсемейства Diplodocoidea, куда также входит группа Flagellicaudata[англ.]. Совместно с сестринской группой, Limaysaurinae[англ.], Nigersaurinae образует кладу Khebbashia[англ.][14].
В описании 2015 года была отмечена эволюционная связь Nigersaurus, Demandasaurus и Tataouinea. В этой системе отсутствует подсемейство Nigersaurinae, вместо него вводится подразделение того же ранга — Rebbachisaurinae[англ.], куда, помимо вышеуказанных родов, входят Katepensaurus[англ.] и Rebbachisaurus[15]. Эта версия получила поддержку авторов описания Lavocatisaurus 2018 года [16].
Ясно, что для Rebbachisauridae короткие шеи и сравнительно небольшие размеры были тенденцией. Пневматизация скелетов членов семейства Rebbachisauridae развивалась постепенно, что наиболее заметно при исследовании костей животных из подсемейства Nigersaurinae[9].
Род Tataouinea является монотипическим. В его состав входит один вид — Tataouinea hannibalis[2].
Кладограмма авторства Фанти и коллег (2013)[14]
| Rebbachisauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Кладограмма авторства Фанти и коллег (2015)[15]
| Rebbachisauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Эволюционное развитие и палеобиогеография

Ещё до открытия Tataouinea отмечалось сходство мегафауны Африки и Европы в раннем мелу. Европейские формы (например, Demandasaurus darvini и Histrisaurus[англ.]) относятся к тому же ярусу, что и Tataouinea; таким образом, члены семейства Rebbachisauridae существовали одновременно по обе стороны океана[12]. Это указывает на то, что данные участки суши были соединены сухопутными мостами[англ.], проходившими через океан Тетис[14][17].
Вероятно, ранние формы Rebbachisauridae появились ещё до разлива Атлантического океана. Так, животные, обитавшие в западной части Гондваны (нынешняя Южная Америка), дали начало подсемейству Limaysaurinae, тогда как мигрировавшие в Африку животные не имели возможности вернуться и обосновались там[18], адаптировавшись для поедания низкорастущих голосеменных растений[19], чтобы не соперничать с более развитыми членами Diplodocoidea и титанозаврами[20].
Океан, затоплявший часть северной Африки, вынуждал динозавров мигрировать к югу[18]. Крупные узкоспециализированные формы, подобные Tataouinea, уменьшались в размерах с уменьшением количества пищи, поэтому более поздняя веха эволюции Rebbachisauridae — Nigersaurus taqueti — была более чем на треть меньше Tataouinea hannibalis[12].
Исчезновение потомков Tataouinea, вероятно, было обусловлено изменением растительности. Не имея из-за узкой специализации возможности конкурировать с высокоразвитыми титанозаврами, Rebbachisauridae были ими вытеснены[20].
Палеобиология и палеоэкология
Будучи растительноядными динозаврами[1], Tataouinea питались наиболее распространенными в то время голосеменными, в том числе и хвойными[21]. Вероятно, динозавры рода Tataouinea питались невысокими растениями, чтобы избежать конкуренции с крупными титанозаврами, лучше приспособленными к поеданию высоко растущих листьев и хвои. Форма зубов Tataouinea указывала на то, что животные просто ощипывали растительность, никак не пережевывая её. Как все зауроподы, Tataouinea, вероятно глотали мелкие камешки — гастролиты — чтобы они, перетирая в кашицу непережёванный растительный материал внутри желудка, помогали усвоению пищи[21]. Существует и другая версия использования гастролитов динозаврами: они глотали их, чтобы получить некоторые минеральные вещества[22].
Динозавр был квадрупедальным (четвероногим). В связи с тем, что передние ноги были короче, бёдра динозавра поднимались над холкой. Хвост был крайне длинным; вероятно, как и у других родственных диплодоку динозавров, он использовался как оружие[23].
Часто, изучая окаменелости зауропод, палеонтологи обнаруживают ископаемые остатки нескольких особей на довольно маленькой площади, это косвенно указывает на стадность этих динозавров. Взрослые, таким образом, могли испытывать меньше затруднений с поиском полового партнёра, а детёныши — рассчитывать на защиту старших[24]. В случае с Tataouinea, однако, известны остатки всего одной особи, так что о стадности динозавров данного рода пока что можно лишь строить предположения[1].
Известные остатки Tataouinea были обнаружены в песчаниках формации Айн-эль-Гуттер, в части Ум-эд-Диаб, датированной ранним альбским веком (возрастом 110 млн лет)[1]. В Ум-эд-Диаб чередуются мелководные, литоральные и неморские отложения[25], образовавшиеся на прибрежной низменности с редкой растительностью и песчаном побережье, омываемом волнами недавно образовавшегося Атлантического океана[1]. Прилежащая область Шенини[англ.] (англ. Chenini member) была влажной и болотистой с мангровыми лесами и разветвлёнными речными системами[26].
Так как почва была маломощна и состояла в основном из измельчённой горной породы, песка и глины, а уровень подземных вод был довольно высок, что следует из состояния корней окаменелых растений[1], маловероятно, что в областях Шенини и Ум-эд-Диаб хватало питания для таких крупных растительноядных, как Tataouinea. Из этого следует вывод, что пастбища Tataouinea находились в другой части Северной Африки, а через Ум-эд-Диаб проходил один из миграционных маршрутов[27].
В реках и прибрежных отмелях обитали небольшие акулы (Lissodus, Leptostyrax[англ.], Scapanorhynchus), двоякодышащие рыбы (Retodus[англ.]) и пилорылые скаты Onchopristis. В тех же отложениях находили ископаемые остатки лучеперых и лопастеперых рыб. Берега населяли гигантские аллигаторы Sarcosuchus, а также мелкие крокодилоподобные животные, в том числе Hamadasuchus[англ.] и Araripesuchus. Также в Ум-эд-Диаб находили остатки птерозавров[1][25], в Шенини среди таких находок — представитель клады Ornithocheiroidea[англ.], вероятно, прилетавший ловить рыбу в местных реках[26].
Многочисленные кости динозавров, обнаруженные в Ум-эд-Диаб и Шенини, принадлежали завроподам-титанозаврам, орнитоподам и тероподам, в частности неизвестным абелизаврам, Carcharodontosaurus и Spinosaurus[1][25][28].
Примечания
- ↑ 1 2 3 4 5 6 7 8 9 10 11 Fanti et al., 2013, Introduction.
- ↑ 1 2 3 Fanti et al., 2013, Abstract.
- ↑ 1 2 Fanti et al., 2013, Fig. 3.
- ↑ Fanti et al., 2013, Etymology.
- ↑ Fanti et al., 2015.
- ↑ 1 2 Paul, 2016, p. 208.
- ↑ 1 2 Paul, 2016, p. 209.
- ↑ 1 2 3 4 5 Sereno et al., 2007, Skeletal Reconstruction.
- ↑ 1 2 Fanti et al., 2013, Osteological correlates of pelvic pneumatization.
- ↑ Fanti et al., 2013, Table 1:Sacrum and total body lengths of selected diplodocoids.
- ↑ Fanti et al., 2013, Diagnosis.
- ↑ 1 2 3 4 Fanti et al., 2013, Discussion.
- ↑ Wedel M. J. Vertebral Pneumaticity, Air Sacs, and the Physiology of Sauropod Dinosaurs (англ.) // Paleobiology. — 2003. — Vol. 29, no. 2. — P. 243–255.
- ↑ 1 2 3 Fanti et al., 2013, Phylogenetic analysis.
- ↑ 1 2 Fanti et al., 2015, Phylogenetic analysis.
- ↑ Canudo et al., 2018, Phylogenetic analysis.
- ↑ Sereno et al., 2007, Diplodocoid Phylogeny.
- ↑ 1 2 Canudo et al., 2018, Paleobiogeographical remarks.
- ↑ Sereno et al., 2007, Feeding Function.
- ↑ 1 2 Fanti, Cau, Hassine, 2014.
- ↑ 1 2 Sander et al., 2011.
- ↑ Wings & Sander, 2007.
- ↑ Upchurch, Barret & Dodson, 2004, Paleobiology, Taphonomy, and Paleoecology.
- ↑ Upchurch, Barret & Dodson, 2004, Examples of Dinosaur Taphonomy Studies.
- ↑ 1 2 3 Fanti et al., 2015, Geological Setting.
- ↑ 1 2 Benton et al., 2000.
- ↑ Fanti et al., 2015, Facies analysis and paleoecology.
- ↑ Upchurch, Barret & Dodson, 2004.
Литература
- Benton M. J., Bouaziz S., Buffetaut E., Martill D., Ouaja M., Soussi M., Trueman C. Dinosaurs and other fossil vertebrates from fluvial deposits in the Lower Cretaceous of southern Tunisia (англ.) // Palaeogeography, Palaeoclimatology, Palaeoecology : журнал. — Amsterdam: Elsevier, 2000. — April (vol. 157, iss. 3—4). — P. 227–246. — doi:10.1016/s0031-0182(99)00167-4.
- Canudo J., Carballido J., Garrido A., Salgado L. A new rebbachisaurid sauropod from the Aptian–Albian, Lower Cretaceous Rayoso Formation, Neuquén, Argentina (англ.) // Acta Palaeontologica Polonica : журнал. — Институт Палеобиологии,Польская Академия Наук, 2018. — Vol. 63, iss. 4. — P. 679–691. — ISSN 0567-7920. — doi:10.4202/app.00524.2018.
- Fanti F., Cau A., Cantelli L., Hassine M., Auditore M. New Information on Tataouinea hannibalis from the Early Cretaceous of Tunisia and Implications for the Tempo and Mode of Rebbachisaurid Sauropod Evolution (англ.) // PLoS One : журнал / Frake A. A.. — Public Library of Science, 2015. — 29 May (vol. 10, iss. 4). — P. e0123475. — doi:10.1371/journal.pone.0123475. — PMID 25923211.
- Fanti F., Cau A., Hassine M. Evidence of titanosauriforms and rebbachisaurids (Dinosauria: Sauropoda) from the Early Cretaceous of Tunisia (англ.) // Journal of African Earth Sciences : журнал. — 2014. — October (no. 90). — P. 1—8. — doi:10.1016/j.jafrearsci.2013.10.010.
- Fanti F., Cau A., Hassine M. & Contessi M. A new sauropod dinosaur from the Early Cretaceous of Tunisia with extreme avian-like pneumatization (англ.) // Nature Communications. — London: Nature Publishing Group, 2013. — Vol. 4, iss. 2080. — P. 1–7. — doi:10.1038/ncomms3080. — . — PMID 23836048.
- Paul G. S. Sauropods // The Princeton field guide to dinosaurs (англ.). — 2nd. — Princeton University press, 2016. — P. 207—209. — 360 p. — (Princeton field guides). — ISBN 978-0-691-16766-4.
- Sander P. M., Christian A., Clauss M., Fechner R., Gee C. T., Griebeler E.-M., Gunga H.-C., Hummel J., Mallison H., Perry S. F., Preuschoft H., Rauhut O. W. M., Remes K., Tütken T., Wings O., Witzel U. Biology of the sauropod dinosaurs: the evolution of gigantism (англ.) // Biological Reviews of the Cambridge Philosophical Society : журнал. — Сambridge: Cambridge University Press, 2011. — February (vol. 86, iss. 1). — P. 117–155. — doi:10.1111/j.1469-185X.2010.00137.x. — PMID 21251189.
- Sereno P. C., Wilson J. A., Witmer L. M., Whitlock J. A., Maga A., Ide O., Rowe T. A. Structural Extremes in a Cretaceous Dinosaur (англ.) // PLoS One : журнал / Frake A. A.. — Public Library of Science, 2007. — 29 November (vol. 2, iss. 11). — P. e0123475. — doi:10.1371/journal.pone.0001230. — . — PMID 18030355.
- Upchurch P., Barrett P. M., Dodson P. Sauropoda (англ.) // Weishampel D. B., Dodson P., Osmólska H. The Dinosauria (2nd Edition) : сборник статей. — University of California Press, 2004. — P. 273—322. — ISBN 9780520941434.
- Whitlock J. A. Inferences of Diplodocoid (Sauropoda: Dinosauria) Feeding Behavior from Snout Shape and Microwear Analyses (англ.) // PloS ONE : журнал / Frake A. A.. — PLOS, 2011. — 6 April (vol. 6, iss. 4). — P. e18304. — doi:10.1371/journal.pone.0018304. — . — PMID 21494685.
- Wings O., Sander M. P. No gastric mill in sauropod dinosaurs: new evidence from analysis of gastrolith mass and function in ostriches (англ.) // Proceedings of the Royal Society B : журнал. — Royal Society Publishing, 2007. — 7 March (vol. 274, iss. 1640). — P. 635–640. — doi:10.1098/rspb.2006.3763. — PMID 17254987.
Ссылки
- Tataouinea (англ.). Prehistoric Wildlife. prehistoric-wildlife.com.