
Митохо́ндрия — двумембранная сферическая или эллипсоидная органелла диаметром обычно около 1 микрометра. Характерна для большинства эукариотических клеток, как автотрофов, так и гетеротрофов. Энергетическая станция клетки; основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии для генерации электрического потенциала, синтеза АТФ и термогенеза. Эти три процесса осуществляются за счёт движения электронов по электронно-транспортной цепи белков внутренней мембраны. Количество митохондрий в клетках различных организмов существенно отличается: так, одноклеточные зелёные водоросли и трипаносомы имеют лишь одну гигантскую митохондрию, тогда как ооцит и амёба Chaos chaos содержат 300 000 и 500 000 митохондрий соответственно; у кишечных анаэробных энтамёб и некоторых других паразитических простейших митохондрии отсутствуют. В специализированных клетках органов животных содержатся сотни и даже тысячи митохондрий.

Фотоси́нтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов.

Динофлагелля́ты, или динофи́товые во́доросли, или динофи́ты, или перидине́и, или па́нцирные жгутиконо́сцы — крупная группа протистов из надтипа альвеолят (Alveolata), которой традиционно присваивают ранг типа. Известно около 4000 ископаемых и более 2500 современных видов, из которых 90 % обитает в морях, остальные — в пресных водах. Около половины представителей — свободноживущие фотосинтезирующие организмы, однако известны и бесцветные гетеротрофные формы, и паразитические динофлагелляты. Некоторые виды являются симбионтами коралловых полипов и двустворчатых моллюсков.

Аденозинтрифосфатсинта́за (АТФ-синта́за, АТФ-фосфогидролаза, H+-transporting two-sector ATPase) — группа ферментов, относящихся к классу транслоказ и синтезирующих аденозинтрифосфат (АТФ) из аденозиндифосфата (АДФ) и неорганического фосфата. Название по номенклатуре — АТФ-фосфогидролаза, однако с августа 2018 года фермент перенесён из третьего (3.6.3.14) в седьмой класс (7.1.2.2), так как катализируемая ферментом реакция протекает по пути, противоположному гидролизу, и не может быть описана с помощью других типов реакций, характеризующие прочие классы ферментов.
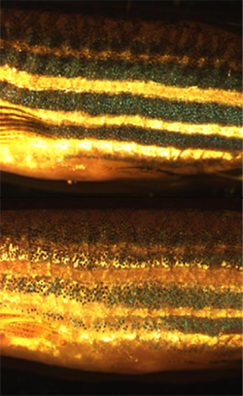
Хроматофоры — пигментсодержащие и светоотражающие клетки, присутствующие у земноводных, рыб, рептилий, ракообразных и головоногих. Они отвечают за окраску кожи и глаз у холоднокровных животных и рождаются в нервном гребне во время эмбриогенеза. Созревшие хроматофоры разделяются на подклассы по цвету в белом свете:

Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.
Вторичные посредники, или «вторичные мессенджеры» — это внутриклеточные сигнальные молекулы, высвобождаемые в тех или иных внутриклеточных сигнальных каскадах в ответ на стимуляцию тех или иных рецепторов и вызванную ею активацию первичных эффекторных белков. Вторичные посредники, в свою очередь, приводят к активации вторичных эффекторных белков. Это, в свою очередь, запускает каскад тех или иных физиологических изменений, которые могут быть важны для обеспечения таких важных физиологических процессов, как рост, развитие и дифференцировка клеток, активация деления клетки, транскрипция или, наоборот, угнетение транскрипции тех или иных генов, биосинтез тех или иных белков, выделение ею гормонов, нейромедиаторов или цитокинов соответственно типу клетки, изменение биоэлектрической активности клетки, миграция клеток, обеспечение их выживаемости или, наоборот, индукция апоптоза. Вторичные посредники являются инициирующими элементами во множестве внутриклеточных сигнальных каскадов. Вследствие всего этого вторичные посредники играют очень важную роль в жизни клетки, а грубое нарушение работы любой из систем вторичных посредников оказывает неблагоприятное воздействие на клетку.
Вестерн-блоттинг — аналитический метод, используемый для определения в образце специфичных белков. На первом этапе используют электрофорез белков в полиакриламидном геле для разделения денатурированных полипептидов по длине или по трехмерной структуре белка. Далее белки переносят на нитроцеллюлозную или PVDF-мембрану, затем детектируют с использованием антител, специфичных к заданному белку.

Глициновый рецептор — один из наиболее распространенных тормозных рецепторов в центральной нервной системе, белок-рецептор с четвертичной структурой, находящийся на постсинаптической мембране многих нейронов, чьим лигандом является глицин. Глициновый рецептор играет важную роль в передаче тормозного сигнала в ЦНС.

Фикобили́ны — тетрапиррольные пигменты (билины) красных водорослей, криптофит и цианобактерий. Фикобилины являются хромофорной группой фикобилипротеинов — кислых водорастворимых глобулярных хромопротеинов светособирающего комплекса водорослей. Отдельными молекулами фикобилины, как правило, не представлены, а образуют комплексы с белками — фикобилипротеиды (хромопротеиды).

Циане́ллы — пластиды глаукофитовых водорослей. Окрашены в сине-зелёный цвет, так как зелёный хлорофилл a маскируется пигментами фикоцианином и аллофикоцианином, расположенными на поверхности тилакоидов. Особое название используется для них потому, что они обладают уникальным признаком: имеют тонкую клеточную стенку из пептидогликана (муреина) между наружной и внутренней мембраной. По большинству других признаков цианеллы напоминают типичные цианобактерии, хотя содержат сильно редуцированный геном.
Холерный токсин — мультисубъединичный белковый токсин, вырабатываемый холерным вибрионом. Вибрион (вирулентный штамм Vibrio cholerae[1] [2] секретирует ХТ после попадания бактерии в организм человека. Действие ХТ является причиной интенсивного обезвоживания после начала активной фазы холерной инфекции. При проникновении в клетки заражённого организма одна из субъединиц этого токсина катализирует АДФ-рибозилирование Gsα-компонента аденилатциклазы, что ведёт к её гиперактивации. Повышенная активность адениталциклазы приводит к нарушению транспорта ионов через мембрану клеток кишечника: снижается поступление ионов натрия и увеличивается отток анионов. В результате происходит быстрая потеря воды клетками кишечника, которая в некоторых случаях доходит до двух литров в час.

Фикобилисо́мы — светособирающие органеллы для фотосистемы II у цианобактерий, красных водорослей и глаукофитов. Стандартные фикобилисомы отсутствуют у криптофитовых и тех представителей прохлорофитовых, у которых имеются фикобилипротеины. У криптофитовых фикобилипротеины находятся во внутритилакоидном пространстве.

Фикоциани́н — это пигмент-белковый комплекс из семейства светособирающих фикобилипротеинов, в которую также входят аллофикоцианин и фикоэритрин. Название происходит от греч. phyco — «водоросль» и cyanin от англ. cyan, что изначально означает оттенок зелёно-голубого и происходит из греч. kyanos, обозначавшего другой цвет — тёмно-голубой. Он является вспомогательным пигментом хлорофилла. Все фикобилипротеины водорастворимы, так что не могут находиться в мембране как каротиноиды, а вместо этого аггрегируются в оформленные кластеры, которые прилипают к мембране фикобилисом. Фикоцианин светло-голубого цвета, поглощает оранжевый и красный свет около 620 нм и излучает в районе 650 нм.
Фикоэритри́н — дополнительный красный фотосинтетический пигмент, обнаруженный у цианобактерий и красных водорослей. Фикоэритрин, как и все фикобилипротеины состоит из белковой части, построенной из α- и β- частиц, организованных в виде шестиугольника и ковалентно связанной с хромофорами, называемыми фикобилинами. Фикоэритрины способны связываться с наибольшим количеством фикобилинов.

Протохлорофиллид, или моновинил протохлорофиллид, это непосредственный предшественник хлорофилла a с отсутствующим фитольным хвостом. В отличие от хлорофилла, протохлорофиллид имеет сильную флюоресценцию; мутанты, накапливающие его в своих тканях, светятся красным, если облучить их синим светом. У цветковых растений, реакция превращения протохлорофиллида в хлорофилл является светозависимой, и такие растения становятся белыми (хлорозными) если выращивать их в темноте. В отличие от них голосеменные, водоросли, и фотосинтезирующие бактерии используют другой, не зависящий от света фермент, и вырастают зелёными даже в темноте.

Светособирающие комплексы — пигмент-белковые комплексы фотосинтезирующих организмов, локализованные в фотосинтетических мембранах и выполняющие функцию первичного поглощения квантов света с последующей миграцией энергии возбуждения к реакционным центрам фотосистем. Также они обеспечивают тонкую настройку фотосинтетического аппарата и участвуют в его защите от фотоповреждений.

Реакционный центр — комплекс белков, пигментов и других кофакторов, взаимодействие которых обеспечивает реакцию превращения энергии света в химическую при фотосинтезе. Реакционный центр получает энергию или через непосредственное возбуждение одной из своих молекул или через перенос энергии от светособирающих комплексов, что даёт начало цепочке химических реакций, происходящей на связанных белками кофакторах. Эти кофакторы — светопоглощающие молекулы такие как хлорофилл, феофитин и хиноны. Энергия фотона используется для поднятия электрона на более высокий энергетический уровень. Запасённая таким образом свободная энергия идёт на восстановление цепочки акцепторов электрона с более высоким редокс-потенциалом.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.
