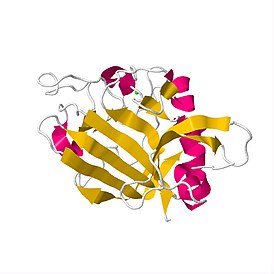
Апопто́з — регулируемый процесс программируемой клеточной гибели, в результате которого клетка распадается на отдельные апоптотические тельца, ограниченные плазматической мембраной. Фрагменты погибшей клетки обычно очень быстро фагоцитируются макрофагами либо соседними клетками, минуя развитие воспалительной реакции. Морфологически регистрируемый процесс апоптоза продолжается 1—3 часа. Одной из основных функций апоптоза является уничтожение дефектных клеток. В многоклеточных организмах апоптоз к тому же задействован в процессах дифференциации и морфогенеза, в поддержании клеточного гомеостаза, в обеспечении важных аспектов развития и функционирования иммунной системы. Апоптоз наблюдается у всех эукариотов, начиная от одноклеточных простейших и вплоть до высших организмов. В программируемой смерти прокариотов участвуют функциональные аналоги эукариотических белков апоптоза.

Систе́ма комплеме́нта — комплекс защитных белков, постоянно присутствующих в крови. Это каскадная система протеолитических ферментов, предназначенная для гуморальной защиты организма от действия чужеродных агентов, она участвует в реализации иммунного ответа организма. Является важным компонентом как врождённого, так и приобретённого иммунитета. Выделяют три основных пути активации системы комплемента: классический, альтернативный и лектиновый. Для запуска классического пути комплемента необходима опсонизация чужеродной клетки антителами, а альтернативный и лектиновый пути могут активироваться в отсутствие антител. Поздние стадии у всех трёх путей активации системы комплемента одинаковы и включают образование мембраноатакующего комплекса, который нарушает целостность мембраны клетки-патогена и приводит к её гибели.

Клеточный рецептор — молекула на поверхности клетки, клеточных органелл или растворенная в цитоплазме. Специфично реагирует изменением своей пространственной конфигурации на присоединение к ней молекулы определённого химического вещества, передающего внешний регуляторный сигнал и, в свою очередь, передает этот сигнал внутрь клетки или клеточной органеллы, нередко при помощи так называемых вторичных посредников или трансмембранных ионных токов.

Кле́тки Ку́пфера — специализированные клетки, располагающиеся в люмене синусоид печени и прикрепленные к синусоидальным эндотелиальным клеткам, образующим стенки кровеносных сосудов. Клетки Купфера — наиболее многочисленные резидентные макрофаги в печени человека, они входят в состав системы мононуклеарных макрофагов. Через воротную вену в печень из пищеварительного тракта поступают кишечные бактерии, бактериальные эндотоксины, которые в печени первым делом сталкиваются с клетками Купфера, образующими первую линию иммунной защиты печени. Нарушения функционирования клеток Купфера сопровождают многие заболевания печени, такие как алкогольная болезнь печени, вирусные гепатиты, внутрипечёночный холестаз, стеатогепатит, цирроз печени, а также при отторжении трансплантата печеночной ткани.

Антагонист в биохимии и фармакологии — подтип лигандов к клеточным рецепторам. Лиганд, обладающий свойствами антагониста рецепторов — это такой лиганд, который блокирует, снижает или предотвращает вызываемые связыванием агониста с рецептором физиологические эффекты. Сам же он при этом не обязан производить какие-либо физиологические эффекты вследствие своего связывания с рецептором. Таким образом, антагонисты рецепторов имеют сродство (аффинность) к данному конкретному типу рецепторов, но, исходя из строгого определения, не имеют собственной внутренней агонистической активности по отношению к этому рецептору, и их связывание лишь нарушает взаимодействие [конкурентных] полных или частичных агонистов с рецептором и предотвращает или ингибирует их функцию и их физиологические эффекты. В равной степени антагонисты рецепторов предотвращают и воздействие на рецептор обратных агонистов. Антагонисты рецепторов опосредуют свои эффекты благодаря связыванию либо с активным сайтом рецептора, тем же самым, с которым связывается и физиологический эндогенный агонист, либо с аллостерическими сайтами, или же они могут взаимодействовать с рецептором в уникальных участках связывания, которые не являются нормальными участками связывания эндогенных веществ для данного рецептора и не принимают в норме участия в физиологической регуляции активности данного рецептора.

Циклический аденозинмонофосфат — органическое соединение, производное АТФ, выполняющее в организме роль вторичного посредника, использующегося для внутриклеточного распространения сигналов некоторых гормонов, которые не могут проходить через клеточную мембрану.
Перици́т, или клетка Руже́ — отростчатая клетка соединительной ткани. Перициты входят в состав стенок мелких кровеносных сосудов, в том числе капилляров. Предшественниками перицитов являются адвентициальные клетки. Впервые перициты описал Шарль Мари Бенджамин Руже в 1874 году. Название «клетка Руже» впервые употребил Циммерман в 1923 году. Эти клетки являются составной частью гематоэнцефалического барьера (ГЭБ). Они обладают несколькими важными для его функционирования свойствами: способностью к сокращению, регулированию функций эндотелия и макрофагальной активностью.

Передача сигнала — в молекулярной биологии термин «Передача сигнала» относится к любому процессу, при помощи которого клетка превращает один тип сигнала или стимула в другой.

Есте́ственные ки́ллеры, также натура́льные ки́ллеры, NK-кле́тки , — тип цитотоксических лимфоцитов, участвующих в функционировании врождённого иммунитета. Функционально NK-клетки аналогичны цитотоксическим T-лимфоцитам (T-киллерам) адаптивного иммунитета позвоночных. NK-клетки обеспечивают ответ на заражение внутриклеточными бактериями и вирусами, уничтожая инфицированные клетки, а также принимают участие в работе противоопухолевого иммунитета. В отличие от других иммунных клеток, NK-клетки распознают заражённые клетки без участия главного комплекса гистосовместимости (MHC) на их мембране, а также антител, благодаря чему ответ, опосредуемый NK-клетками, является очень быстрым.

Толл-подобные рецепторы — класс клеточных рецепторов с одним трансмембранным фрагментом, которые распознают консервативные структуры микроорганизмов и активируют клеточный иммунный ответ. Играют ключевую роль во врождённом иммунитете. Например, толл-подобный рецептор 4 узнаёт и связывается с консервативной структурой клеточной стенки грамотрицательных бактерий — липополисахаридом. Название получили благодаря сходству с белком, который кодируется открытым в 1985 году геном Toll у дрозофилы.
Вторичные посредники, или «вторичные мессенджеры» — это внутриклеточные сигнальные молекулы, высвобождаемые в тех или иных внутриклеточных сигнальных каскадах в ответ на стимуляцию тех или иных рецепторов и вызванную ею активацию первичных эффекторных белков. Вторичные посредники, в свою очередь, приводят к активации вторичных эффекторных белков. Это, в свою очередь, запускает каскад тех или иных физиологических изменений, которые могут быть важны для обеспечения таких важных физиологических процессов, как рост, развитие и дифференцировка клеток, активация деления клетки, транскрипция или, наоборот, угнетение транскрипции тех или иных генов, биосинтез тех или иных белков, выделение ею гормонов, нейромедиаторов или цитокинов соответственно типу клетки, изменение биоэлектрической активности клетки, миграция клеток, обеспечение их выживаемости или, наоборот, индукция апоптоза. Вторичные посредники являются инициирующими элементами во множестве внутриклеточных сигнальных каскадов. Вследствие всего этого вторичные посредники играют очень важную роль в жизни клетки, а грубое нарушение работы любой из систем вторичных посредников оказывает неблагоприятное воздействие на клетку.
Протеинкина́зы — подкласс ферментов киназ (фосфотрансфераз). Протеинкиназы модифицируют другие белки путём фосфорилирования остатков аминокислот, имеющих гидроксильные группы или гетероциклической аминогруппы гистидина.
-селектин — гликопротеин, находящийся на клеточной поверхности, относится к классу молекул клеточной адгезии; рецептор к некоторым углеводным лигандам лейкоцитов крови. Вырабатывается клетками эндотелия в случае воспалительного повреждения ткани, например инфицировании, и способствует рекрутированию нейтрофилов из циркулирующей крови к месту повреждения. Наиболее распространённый и более изученный из 3 селектинов.
Трансмембранные рецепторы — мембранные белки, которые размещаются, и работают не только во внешней клеточной мембране, но и в мембранах компартментов и органелл клетки. Связывание с сигнальной молекулой происходит с одной стороны от мембраны, а клеточный ответ формируется на другой стороне от мембраны. Таким образом, они играют уникальную и важную роль в межклеточных связях и передаче сигнала.
Eph-рецепторы — группа рецепторов, принадлежащих к семейству рецепторов-тирозинкиназ и связывающих эфрины (Eph). Вместе с лигандами участвуют в процессах, отвечающих за эмбриональное развитие организма, например, в сегментации, аксональном наведении, миграции клеток. Также они участвуют и в процессах, происходящих во взрослом организме, таких как долговременная потенциация, ангиогенез, дифференциация стволовых клеток и формирование раковых опухолей. И рецепторы, и лиганды являются мембранными белками и взаимодействуют при непосредственном контакте клеток.

Кореце́птор — находящийся на поверхности клетки дополнительный рецептор, который связывается с сигнальной молекулой (лигандом) в дополнение к первичному рецептору. Корецепторы Т-лимфоцитов усиливают взаимодействие T-клеточного рецептора с комплексом пептид + молекула главного комплекса гистосовместимости (MHC). Корецепторы нередко используются вирусами для проникновения в клетку.

Тирозинкиназа, тирозин-специфичная протеинкиназа — фермент подкласса протеинкиназ, группы киназ (фосфотрансфераз). Тирозинкиназы катализируют перенос фосфатного остатка от АТФ на тирозиновый остаток специфических клеточных белков-мишеней. Тирозинкиназы — одно из важнейших звеньев в системе передачи сигналов в клетке.
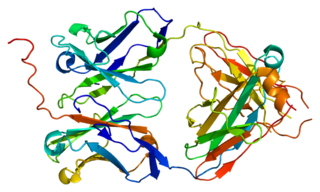
Пре-B-клеточный рецептор — рецепторный белковый комплекс, который присутствует на плазматической мембране предшественников B-клеток в течение короткого времени и служит сигналом завершения продуктивной соматической рекомбинации генов тяжёлых цепей иммуноглобулинов. Хотя пре-В-клеточный рецептор синтезируется в клетке только в течение короткого промежутка времени, это событие, тем не менее, является важной контрольной точкой в развитии В-лимфоцита. Его присутствие на мембране показывает, что клетки способны синтезировать нормальные тяжёлые цепи иммуноглобулинов, и только такие клетки могут продолжать своё развитие, в противном случае они погибают в результате апоптоза. Синтез пре-B-клеточного рецептора знаменует переход от про-B-клеток к пре-B-клеткам.
Рецепторы подтипа 5-HT1A обладают несколькими различными внутриклеточными сигнальными путями.

BMPR2 — рецептор с серин/треонинкиназной ативностью. Он связывает BMP бета-субъединиц TGF-лигандов, вовлеченных в паракринную сигнализацию, что является формой коммуникации клеток, в которой клетка передает сигнал, вызывающий изменения в соседних клетках, изменяя их поведение или дифференцировку. Передача сигналов в BMP-пути начинается с присоединения BMP к рецептору II типа. Это вызывает вовлечение BMP рецептора I типа, который подвергается фосфорелированию. Фосфорилат рецептора I типа — R-SMAD транскрипционный регулятор.