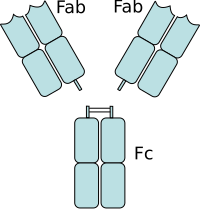
Антитела́, иммуноглобулины — крупные глобулярные белки плазмы крови, выделяющиеся плазматическими клетками иммунной системы и служащие для нейтрализации клеток патогенов и вирусов, а также белковых ядов и некоторых других чужеродных веществ. Каждое антитело распознаёт уникальный элемент патогена, отсутствующий в самом организме, — антиген, а в пределах данного антигена — определённый его участок, эпитоп. Связываясь с антигенами на поверхности патогенов, антитела могут либо непосредственно нейтрализовать их, либо привлекать другие компоненты иммунной системы, такие как система комплемента и фагоциты, чтобы уничтожить чужеродные клетки или вирусные частицы. Антитела — важнейший компонент гуморального специфического иммунитета.

Ферме́нты, или энзи́мы , — обычно сложные белковые соединения, РНК (рибозимы) или их комплексы, ускоряющие химические реакции в живых системах. Каждый фермент, свернутый в определённую структуру, ускоряет соответствующую химическую реакцию: реагенты в такой реакции называются субстратами, а получающиеся вещества — продуктами. Ферменты специфичны к субстратам: АТФ-аза катализирует расщепление только АТФ, а киназа фосфорилазы фосфорилирует только фосфорилазу.

Систе́ма комплеме́нта — комплекс защитных белков, постоянно присутствующих в крови. Это каскадная система протеолитических ферментов, предназначенная для гуморальной защиты организма от действия чужеродных агентов, она участвует в реализации иммунного ответа организма. Является важным компонентом как врождённого, так и приобретённого иммунитета. Выделяют три основных пути активации системы комплемента: классический, альтернативный и лектиновый. Для запуска классического пути комплемента необходима опсонизация чужеродной клетки антителами, а альтернативный и лектиновый пути могут активироваться в отсутствие антител. Поздние стадии у всех трёх путей активации системы комплемента одинаковы и включают образование мембраноатакующего комплекса, который нарушает целостность мембраны клетки-патогена и приводит к её гибели.

T-лимфоциты, или Т-клетки — лимфоциты, развивающиеся у млекопитающих в тимусе из предшественников — претимоцитов, поступающих в него из красного костного мозга. В тимусе T-лимфоциты дифференцируются, приобретая Т-клеточные рецепторы и различные корецепторы. Играют важную роль в приобретённом иммунном ответе. Обеспечивают распознавание и уничтожение клеток, несущих чужеродные антигены, усиливают действие моноцитов, NK-клеток, а также принимают участие в переключении изотипов иммуноглобулинов.

Гликопротеи́ны — двухкомпонентные белки, в которых белковая (пептидная) часть молекулы ковалентно соединена с одной или несколькими группами гетероолигосахаридов. Кроме гликопротеинов существуют также протеогликаны и гликозаминогликаны.
Приобретённый иммунитет — способность организма обезвреживать чужеродные и потенциально опасные микроорганизмы, которые уже попадали в организм ранее. Представляет собой результат работы системы высокоспециализированных клеток (лимфоцитов), расположенных по всему организму. Считается, что система приобретённого иммунитета возникла у челюстноротых позвоночных. Она тесно взаимосвязана с гораздо более древней системой врождённого иммунитета, которая является основным средством защиты от патогенных микроорганизмов у большинства живых существ.

Иммуноферментный анализ — лабораторный иммунологический метод качественного или количественного определения различных низкомолекулярных соединений, макромолекул, вирусов и пр., в основе которого лежит специфическая реакция антиген-антитело. Выявление образовавшегося комплекса проводят с использованием фермента в качестве метки для регистрации сигнала. Теоретические основы ИФА опираются на современную иммунохимию и химическую энзимологию, знание физико-химических закономерностей реакции антиген-антитело, а также на основные принципы аналитической химии.

Т-клеточный рецептор — поверхностный белковый комплекс Т-лимфоцитов, ответственный за распознавание процессированных антигенов, связанных с молекулами главного комплекса гистосовместимости на поверхности антигенпредставляющих клеток. ТКР состоит из двух субъединиц, заякоренных в клеточной мембране, и ассоциирован с мультисубъединичным комплексом CD3. Взаимодействие ТКР с молекулами ГКГ и связанным с ними антигеном ведёт к активации Т-лимфоцитов и является ключевой точкой в запуске иммунного ответа.

Лёгкие цепи иммуноглобулинов — это белковые субъединицы иммуноглобулинов. Молекулы иммуноглобулинов класса G состоят из двух лёгких и двух тяжёлых цепей. Лёгкие цепи иммуноглобулинов имеют молекулярную массу около 28 кДа.

Антиген — любое вещество, которое организм рассматривает как чужеродное или потенциально опасное и против которого организм обычно начинает вырабатывать собственные антитела. Обычно в качестве антигенов выступают белки, однако простые вещества, даже металлы, также могут становиться антигенами в сочетании с собственными белками организма и их модификациями (гаптены).

Кристаллизующийся фрагмент иммуноглобулина — концевая часть молекулы иммуноглобулина, которая взаимодействует с Fc-рецептором на поверхности клетки и с некоторыми белками системы комплемента. Данное свойство позволяет антителам активировать иммунную систему. Fc-участок изотипов IgG, IgA и IgD состоит из двух одинаковых белковых фрагментов, соответственно, второго и третьего константных доменов двух тяжелых цепей; в случае изотипов IgM и IgE Fc содержит три константных домена тяжелых цепей в каждой полипептидной цепочке.
Бело́к A — белок молекулярной массой 40—60 кДа, выделенный с поверхности клеточной стенки золотистого стафилококка. Белок A используется в биохимических исследованиях, так как хорошо связывает многие иммуноглобулины млекопитающих, особенно иммуноглобулины G.
Тяжёлые цепи иммуноглобулинов — это большие полипептидные субъединицы иммуноглобулинов (антител).

V(D)J-рекомбина́ция, или V(D)J-реаранжиро́вка , — механизм соматической рекомбинации ДНК, происходящий на ранних этапах дифференцировки лимфоцитов и приводящий к формированию антиген-распознающих участков антител и Т-клеточного рецептора. Гены иммуноглобулинов и Т-клеточных рецепторов состоят из повторяющихся сегментов, принадлежащих к трём классам: V, D и J. В процессе V(D)J-перестройки генные сегменты, по одному из каждого класса, соединяются вместе. Объединённая последовательность сегментов V(D)J кодирует вариабельные домены каждой из цепей рецептора или антитела.

Плазматические клетки, плазмоциты — основные клетки, продуцирующие антитела в организме человека. Являются конечным этапом развития B-лимфоцита.
ALCAM — мембранный белок из суперсемейства иммуноглобулинов, относится к классу молекул клеточной адгезии. Связывается с CD6.

Иммуноглобули́ны G (IgG) — класс антител. IgG составляют около 75 % антител плазмы крови у человека и являются наиболее часто встречаемыми в кровотоке антителами. IgG продуцируются плазматическими B-клетками, и каждая молекула IgG имеет два сайта связывания антигена.

Иммуноглобули́ны M (IgM) — класс антител. Молекулы IgM являются самыми тяжёлыми и наиболее сложно организованными иммуноглобулинами. Молекула свободного IgM представляет собой пентамер, каждый мономер которого состоит из двух тяжёлых цепей (μ-цепей) и двух лёгких цепей κ- или λ-типов. Мономеры объединены в пентамер посредством дисульфидных мостиков и J-цепи. При первичном попадании в организм антигена иммуноглобулины IgM из всех антител образуются первыми. Кроме того, они первыми появляются в онто- и филогенезе. У человека и других млекопитающих IgM синтезируются плазматическими клетками, находящимися в селезёнке. IgM наиболее активны в антибактериальном иммунитете и при ряде аутоиммунных заболеваний.
Иммуноглобули́ны Y (IgY) — класс антител, имеющийся у птиц, рептилий и амфибий. IgY накапливаются в большом количестве в желтке яиц, куда транспортируются из плазмы крови. Молекула IgY состоит из двух тяжёлых и двух лёгких цепей. В ранних источниках IgY ошибочно именуют IgG из-за структурного и функционального сходства с иммуноглобулинами G млекопитающих. Однако между IgY и IgG всё же имеются существенные структурные и функциональные различия, а антитела, направленные против IgG (анти-IgG), не действуют на IgY.

Класси́ческий пу́ть актива́ции систе́мы комплеме́нта — один из трёх путей активации системы комплемента, наряду с альтернативным путём и лектиновым путём. Классический путь запускают комплексы антиген-антитело, причём антитела должны принадлежать к классу IgG или IgM. После активации происходит сборка белкового комплекса C3-конвертазы (C4b2b), который разрезает белок комплемента C3. Один из фрагментов C3, C3b, связывается с C3-конвертазой, в результате чего образуется C5-конвертаза (C4b2b3b). C5-конвертаза разрезает компонент комплемента 5, после чего образовавшиеся белковые фрагменты привлекают фагоциты в очаг инфекции и способствуют поглощению клеток патогенов. C5-конвертаза также запускает финальные стадии каскада комплемента, которые завершаются формированием мембраноатакующего комплекса. Он формирует поры в мембране клетки микроорганизма, вызывая её лизис и гибель. Помимо микробных клеток, классический путь комплемента могут запускать апоптотические тельца и некротизированные клетки.