Митохо́ндрия — двумембранная сферическая или эллипсоидная органелла диаметром обычно около 1 микрометра. Характерна для большинства эукариотических клеток, как автотрофов, так и гетеротрофов. Энергетическая станция клетки; основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии для генерации электрического потенциала, синтеза АТФ и термогенеза. Эти три процесса осуществляются за счёт движения электронов по электронно-транспортной цепи белков внутренней мембраны. Количество митохондрий в клетках различных организмов существенно отличается: так, одноклеточные зелёные водоросли и трипаносомы имеют лишь одну гигантскую митохондрию, тогда как ооцит и амёба Chaos chaos содержат 300 000 и 500 000 митохондрий соответственно; у кишечных анаэробных энтамёб и некоторых других паразитических простейших митохондрии отсутствуют. В специализированных клетках органов животных содержатся сотни и даже тысячи митохондрий.

Кле́точная мембра́на — эластическая молекулярная структура, состоящая из белков и липидов. Отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулирует обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определённые условия среды.

Фотоси́нтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов.

Halobacteriaceae (лат.) — семейство архей из класса галобактерий (Halobacteria), единственное в порядке Halobacteriales. Включает около двадцати родов, в том числе Halobacterium, Halococcus, Haloarcula, Natrococcus, Natrobacterium.

Электропорация — создание пор в бислойной липидной мембране под действием электрического поля. Это явление используется в биотехнологии для внедрения макромолекул в клетки млекопитающих, бактерий или растений, а также используется в медицине и промышленности.

Аденозинтрифосфатсинта́за (АТФ-синта́за, АТФ-фосфогидролаза, H+-transporting two-sector ATPase) — группа ферментов, относящихся к классу транслоказ и синтезирующих аденозинтрифосфат (АТФ) из аденозиндифосфата (АДФ) и неорганического фосфата. Название по номенклатуре — АТФ-фосфогидролаза, однако с августа 2018 года фермент перенесён из третьего (3.6.3.14) в седьмой класс (7.1.2.2), так как катализируемая ферментом реакция протекает по пути, противоположному гидролизу, и не может быть описана с помощью других типов реакций, характеризующие прочие классы ферментов.
Мембранный транспорт — транспорт веществ сквозь клеточную мембрану в клетку или из клетки, осуществляемый с помощью различных механизмов — простой диффузии, облегченной диффузии и активного транспорта.

К мембранным белкам относятся белки, которые встроены в клеточную мембрану или мембрану клеточной органеллы или ассоциированы с таковой. Около 25 % всех белков являются мембранными.
Протонный насос в молекулярной биологии — механизм, создающий или использующий протонный градиент. Протонный насос является белковым комплексом, в котором имеются мембранные белки. В процессе работы протонного насоса осуществляется перенос протонов через мембрану клетки, митохондрии или другого внутриклеточного компартмента. Первичными протонными насосами называют комплексы, в которых разность концентраций используется для синтеза АТФ; вторичные протонные насосы расщепляют АТФ для создания протонного градиента.

Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.
Рецепторы, сопряжённые с G-белком,, также известные как семиспиральные рецепторы или серпентины, составляют большое семейство трансмембранных рецепторов. GPCR выполняют функцию активаторов внутриклеточных путей передачи сигнала, приводящими в итоге к клеточному ответу. Рецепторы этого семейства обнаружены только в клетках эукариот: у дрожжей, растений, хоанофлагеллят и животных. Эндогенные лиганды-агонисты, которые связываются и активируют эти рецепторы, включают гормоны, нейромедиаторы, светочувствительные вещества, пахучие вещества, феромоны и варьируются в своих размерах от небольших молекул и пептидов до белков. Нарушение работы GPCR приводит к возникновению множества различных заболеваний, а сами рецепторы являются мишенью до 40 % выпускаемых лекарств. Точный размер надсемейства GPCR не известен, но почти 800 различных человеческих генов были предсказаны из анализа последовательности генома. Несмотря на многочисленные схемы, было предложено разделить надсемейство на три основных класса.
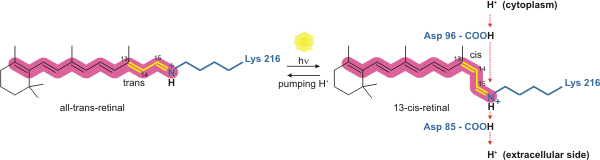
Опсины — это группа светочувствительных связанных с мембраной рецепторов, связанных с G-белками, массой около 35-55 кДа семейства ретинолидных белков, которые обнаружены в фоточувствительных клетках сетчатки. Пять групп опсинов принимают участие в зрении, передаче света в электрохимический сигнал, и являются первым этапом в каскаде зрительной трансдукции. Меланопсин — другой опсин, обнаруженный в сетчатке млекопитающих, который участвует в циркадианных ритмах и в зрачковом рефлексе, но не в формировании изображения.
Бактериородопси́нсодержащие плёнки — наноплёнки, полученные с использованием пурпурных мембран галобактерий, содержащих бактериородопсин. Используются в качестве компонента в биомолекулярной электронике.
Хемиосмос — биохимический механизм, с помощью которого осуществляется превращение энергии цепи переноса электронов в энергию АТФ. Включает изменение электрохимического потенциала клеточной мембраны.

Реакционный центр — комплекс белков, пигментов и других кофакторов, взаимодействие которых обеспечивает реакцию превращения энергии света в химическую при фотосинтезе. Реакционный центр получает энергию или через непосредственное возбуждение одной из своих молекул или через перенос энергии от светособирающих комплексов, что даёт начало цепочке химических реакций, происходящей на связанных белками кофакторах. Эти кофакторы — светопоглощающие молекулы такие как хлорофилл, феофитин и хиноны. Энергия фотона используется для поднятия электрона на более высокий энергетический уровень. Запасённая таким образом свободная энергия идёт на восстановление цепочки акцепторов электрона с более высоким редокс-потенциалом.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.
Интегральный мембранный белок — один из типов мембранных белков, которые прочно связаны с цитоплазматической мембраной (интегрированы). Они составляют значительную часть белков, закодированных в геноме любого организма. Интегральные белки могут быть погружены в мембрану полностью, а иногда даже пронизывают её насквозь. В этом смысле, все трансмембранные белки являются интегральными белками, но не все интегральные — трансмембранными. Связь интегральных белков с мембранными липидами очень прочна и определяется главным образом гидрофобными взаимодействиями.

Электрохими́ческий градие́нт, или градиéнт электрохимического потенциáла, — совокупность градиента концентрации и мембранного потенциала, которая определяет направление движения ионов через мембрану. Состоит из двух составляющих: химического градиента, или разницы в концентрациях растворённого вещества по обе стороны мембраны, и электрического градиента, или разницы зарядов, расположенных на противоположных сторонах мембраны. Градиент возникает вследствие неодинаковой концентрации ионов на противоположных сторонах водопроницаемой мембраны. Ионы двигаются через мембрану из области, имеющую более высокую концентрацию в область с более низкой концентрацией путём простой диффузии. Также ионы несут электрический заряд, который формирует электрический потенциал на мембране. Если существует неравномерное распределение зарядов по обе стороны мембраны, то разница в электрическом потенциале порождает силу, которая приводит к ионной диффузии, пока заряды по обе стороны не будут сбалансированы.
Га́зовые вези́кулы — заполненные газом микрокомпартменты, обеспечивающие повышенную плавучесть клеток у примерно 150 видов планктонных бактерий и архей. Особенно часто газовые вакуоли встречаются у цианобактерий. Мембрана, ограничивающая газовую везикулу, имеет белковую природу, а внутри газовой везикулы находится атмосферный воздух. Газовые везикулы располагаются в цитоплазме поодиночке или образуют сотовидные скопления, которые иногда неправильно называют газовыми вакуолями.