
Гисто́ны — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5, H2A, H2B, H3, H4. Гистоны H2A, H2B, H3, H4, называемые кóровыми гистонами, формируют нуклеосому, представляющую собой белковую глобулу, вокруг которой накручена нить ДНК. Гистон H1/H5, называемый линкерным гистоном, связывается с внешней стороной нуклеосомы, фиксируя на ней нить ДНК. В хроматине гистоны составляют 25—40 % сухого веса. Благодаря высокому содержанию лизина и аргинина гистоны проявляют сильно оснóвные свойства. Гистоны непосредственно контактируют с ДНК и способны нейтрализовать отрицательный заряд фосфатных групп ДНК за счёт положительных зарядов аминокислотных остатков. Последовательность аминокислот в этих белках является консервативной и практически не различается в организмах различных таксонов. Гистоны присутствуют в ядрах эукариотических клеток; у бактерий гистонов нет, но они выявлены у архей группы Euryarchaea.
Нуклеосома — это структурная часть хромосомы, образованная совместной упаковкой нити ДНК с гистоновыми белками H2А, H2B, H3 и H4. Последовательность нуклеосом, соединенная гистоновым белком H1, формирует нуклеофиламент, или иначе нуклеосомную нить.
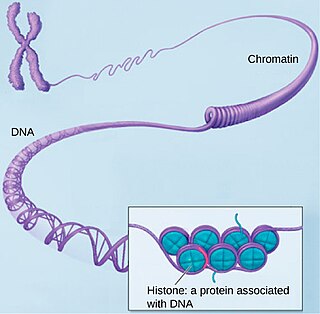
Хроматин — нуклеопротеид, составляющий основу хромосом. Состоит из ДНК и белков. Хроматин находится внутри ядра клеток эукариот и архей, имеющих гистоны. В более широком смысле хроматином иногда называют также и вещество нуклеоида у бактерий.
Гистонацетилтрансфераза (англ. Histone acetyltransferases, HAT) — фермент (КФ 2.3.1.48), из семейства ацилтрансфераз (класс трансферазы), катализирующий реакцию переноса ацетильной группы CH3CO- (реакция ацетилирования) на остатки лизина в гистонах. Донором ацетильной группы является ацетилкофермент А, при этом образуется ε-N-ацетиллизин.

Гетерохромати́н — участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость.

Xist — ген, кодирующий РНК и локализованный на Х-хромосоме плацентарных млекопитающих, является ключевым эффектором в инактивации Х-хромосомы. Он входит в состав комплекса Xic, наряду с двумя другими РНК-кодирующими генами и двумя белоккодирующими генами. Продукт гена Xist — Xist-РНК — представляет собой крупный транскрипт, который экспрессируется на неактивной хромосоме и не экспрессируется на активной. Процессинг этого транскрипта напоминает процессинг мРНК и тоже включает этапы сплайсинга и полиаденилирования, однако он остаётся в ядре и не транслируется. Было высказано предположение, что ген Xist по крайней мере частично образовался как часть белоккодирующего гена, который впоследствии стал псевдогеном. Инактивированная Х-хромосома покрыта Xist-РНК, который необходим для процесса инактивации. Х-хромосома, лишённая гена Xist, не будет инактивирована, однако дупликация этого гена на другой хромосоме вызывает инактивацию и первой хромосомы тоже.
Белки группы polycomb — это семейство белков, которые способны ремоделировать хроматин. Эти белки-регуляторы были впервые описаны у дрозофил, где они подавляют гомеозисные гены, контролирующие индивидуальные отличия сегментов развивающегося эмбриона.

Эухромати́н, также активный или «открытый хромати́н» — участки хроматина, которые представляет собой неплотную и легко упакованную форму, обогащённую генами и часто находящуюся в состоянии активной транскрипции. Эухроматин отличается от гетерохроматина, который плотно упакован и менее доступен для транскрипции. Около 92 % генома человека является эухроматичным.
Эпигенетика — раздел генетики. Эпигенетика изучает наследуемые изменения активности генов во время роста и деления клеток — изменения синтеза белков, вызванных механизмами, не изменяющими последовательность нуклеотидов в ДНК. Эпигенетические изменения сохраняются в ряде митотических делений соматических клеток, а также могут передаваться следующим поколениям. Регуляторы синтеза белка — метилирование и деметилирование ДНК, ацетилирование и деацетилирование гистонов, фосфорилирование и дефосфорилирование транскрипционных факторов и другие внутриклеточные механизмы.

Откры́тый хромати́н — небольшие участки хроматина, свободные от нуклеосом. Посадке нуклеосом, как правило, препятствуют связанные с хроматином белковые факторы, узнающие определённые последовательности ДНК. К числу таких белков относятся транскрипционные факторы, ДНК- или РНК-полимеразы. Открытый хроматин часто совпадает с цис-регуляторными последовательностями, а именно: промоторами, энхансерами, инсуляторами, сайленсерами, участками начала репликации ДНК. Размер открытых участков хроматина обычно составляет несколько сотен пар нуклеотидов, в среднем около 300 п.н.
Позициони́рование нуклеосо́м — определение положения нуклеосом на последовательности ДНК эукариот. Участки ДНК могут либо входить в нуклеосомные комплексы, либо быть в составе линкерной, межнуклеосомной ДНК. Эта характеристика ДНК определяет её доступность для взаимодействия с белками.
Сиртуины — семейство эволюционно консервативных НАД-зависимых белков, обладающих деацетилазной или АДФ-рибозилтрансферазной активностью. Название семейству дано в честь одного из представителей – дрожжевого белка SIR2. Сиртуины обнаружены у многих живых организмов, от бактерий до млекопитающих, и вовлечены в регуляцию важных клеточных процессов и метаболических путей.

Бромодоменсодержащий белок 4 — хроматинсвязывающий белок, который у человека кодируется геном BRD4. Он узнаёт и связывается с остатками ацетилированного лизина в составе гистонов и других белков. BRD4 принимает участие в регуляции транскрипции и передаче эпигенетической информации при делении клетки. В 2016 году было установлено, что BRD4 обладает собственной гистонацетилтрансферазной активностью.
Семейство белков BET — семейство белков, содержащих два тандемных бромодомена на N-конце молекулы и экстратерминальный домен. Белки семейства BET являются глобальными корегуляторами транскрипции. Они связывают ε-аминоацетильные группы остатков лизина в гистонах и привлекают в этот комплекс белки ремоделирования хроматина, соответственно они могут выступать в роли коактиваторов или корепрессоров транскрипции в зависимости от молекулярного контекста. Бромодомены белков семейства BET характеризуются относительно низкой аффинностью к одиночным остаткам ацетиллизина к полипептидах и гораздо большей к участкам со множественными сайтами ацетилирования.

CREB-свя́зывающий бело́к — белок, который у человека кодируется геном CREBBP, локализованным на 16-й хромосоме. CBP функционирует как транскрипционный коактиватор, то есть посредством белок-белковых взаимодействий связывает различные белковые трансактиваторы транскрипции с основным транскрипционным комплексом. Впервые описан исследователями в 1993 году.
Гистондеацетила́за 4 — белок, кодируемый у человека геном HDAC4, расположенным на 2-й хромосоме. Как и все ферменты группы гистондеацетилаз, близкой к сиртуинам, гистондеацетилаза 4 катализирует удаление ацетильных групп с остатков лизина в N-концевой части коровых гистонов, что изменяет структуру хроматина. Деацетилирование гистонов является одним из механизмов транскрипционной и эпигенетической регуляции, оказывает влияние на ход клеточного цикла и участвует в регуляции развития. Работа HDAC4 регулируется путём различных посттрансляционных модификаций и взаимодействий с разнообразными белками, иногда тканеспецифичными. Нарушение работы HDAC4 приводит к развитию многих заболеваний, в том числе раковых, поэтому ингибиторы HDAC4 могут иметь важное медицинское применение.
Нуклеофозми́н — ядрышковый белок, у человека кодируется геном NPM1, локализованным на 5-й хромосоме. Нуклеофозмин перемещается между ядром и цитоплазмой и действует как многофункциональный шаперон нуклеиновых кислот, принимающий участие в таких процессах, как биогенез рибосом, ремоделирование хроматина, регуляция митоза, поддержание стабильности генома, репарация ДНК и транскрипция. Нарушения в работе нуклеофозмина могут приводить к развитию злокачественных новообразований и других заболеваний; в частности, мутации, затрагивающие его ген, приводят к развитию острого миелоидного лейкоза.

Хромодомен — белковый домен длиной около 40-50 аминокислотных остатков, обычно встречающийся в белках, связанных с ремоделированием хроматина. Домен является высоко консервативным среди растений и животных и представлен в составе множества различных белков генома. Некоторые хромодомен-содержащие гены имеют изоформы, образованные за счёт альтернативного сплайсинга и не содержат хромодоменов. У млекопитающих хромодомен-содержащие белки участвуют в процессах регуляции генов, связанных с ремоделированием хроматина и образованием участков гетерохроматина. Хромодомен-содержащие белки также связывают метилированные гистоны и присутствуют в составе комплекса РНК-индуцированного сайленсинга генов.
RITS — форма РНК-интерференции, при которой короткие молекулы РНК, такие как малые интерферирующие РНК (siРНК), подавляют транскрипцию гена-мишени. Это часто сопровождается посттрансляционными модификациями хвостов гистонов, а именно метилированием лизина 9 гистона H3 (H3K9me), которое приводят к образованию гетерохроматина в локусе-мишени. Таким образом, RITS участвует в образовании гетерохроматина de novo. Белковый комплекс, который связывается с siРНК и взаимодействует с метилированным остатком лизина 9 гистона Н3, называется комплексом RITS. RITS был открыт у делящихся дрожжей Schizosaccharomyces pombe, и было показано, что он принимает участие в инициации образования гетерохроматина и его поддержании в локусе типа спаривания и в образовании центромеры. В состав комплекса RITS S. pombe входят три белка: белок группы argonaute, содержащий piwi-домен и похожий на РНКазу Н, белок Chp1, содержащий хромодомен, и белок Tas3, взаимодействующий с белками argonaute и с Chp1. Для образования гетерохроматина необходимы, как минимум, белок argonaute и РНК-зависимая РНК-полимераза. Утрата генов, кодирующих эти белки, у S. pombe, приводит к нарушениям в структуре гетерохроматина и функционировании центромер, так как комплекс RITS содержит siРНК, считанную с центромерных повторов. Аномальное функционирование центромер, в свою очередь, приводит к нарушению сегрегации хромосом в митозе, а именно — к появлению «отстающих» хромосом на стадии анафазы.
Эпигено́мика — раздел молекулярной биологии, изучающий совокупность эпигенетических модификаций генетического материала клетки с помощью высокопроизводительных методов. Эпигеномика аналогична геномике и протеомике, которые изучают геном и протеом клетки, соответственно.