
Клеточный рецептор — молекула на поверхности клетки, клеточных органелл или растворенная в цитоплазме. Специфично реагирует изменением своей пространственной конфигурации на присоединение к ней молекулы определённого химического вещества, передающего внешний регуляторный сигнал и, в свою очередь, передает этот сигнал внутрь клетки или клеточной органеллы, нередко при помощи так называемых вторичных посредников или трансмембранных ионных токов.
Агонист — химическое соединение (лиганд), которое при взаимодействии с рецептором изменяет его состояние, приводя к биологическому отклику. Обычные агонисты увеличивают отклик рецептора, обратные агонисты уменьшают его, а антагонисты блокируют действие рецептора.

Антагонист в биохимии и фармакологии — подтип лигандов к клеточным рецепторам. Лиганд, обладающий свойствами антагониста рецепторов — это такой лиганд, который блокирует, снижает или предотвращает вызываемые связыванием агониста с рецептором физиологические эффекты. Сам же он при этом не обязан производить какие-либо физиологические эффекты вследствие своего связывания с рецептором. Таким образом, антагонисты рецепторов имеют сродство (аффинность) к данному конкретному типу рецепторов, но, исходя из строгого определения, не имеют собственной внутренней агонистической активности по отношению к этому рецептору, и их связывание лишь нарушает взаимодействие [конкурентных] полных или частичных агонистов с рецептором и предотвращает или ингибирует их функцию и их физиологические эффекты. В равной степени антагонисты рецепторов предотвращают и воздействие на рецептор обратных агонистов. Антагонисты рецепторов опосредуют свои эффекты благодаря связыванию либо с активным сайтом рецептора, тем же самым, с которым связывается и физиологический эндогенный агонист, либо с аллостерическими сайтами, или же они могут взаимодействовать с рецептором в уникальных участках связывания, которые не являются нормальными участками связывания эндогенных веществ для данного рецептора и не принимают в норме участия в физиологической регуляции активности данного рецептора.
Рецепторы, сопряжённые с G-белком,, также известные как семиспиральные рецепторы или серпентины, составляют большое семейство трансмембранных рецепторов. GPCR выполняют функцию активаторов внутриклеточных путей передачи сигнала, приводящими в итоге к клеточному ответу. Рецепторы этого семейства обнаружены только в клетках эукариот: у дрожжей, растений, хоанофлагеллят и животных. Эндогенные лиганды-агонисты, которые связываются и активируют эти рецепторы, включают гормоны, нейромедиаторы, светочувствительные вещества, пахучие вещества, феромоны и варьируются в своих размерах от небольших молекул и пептидов до белков. Нарушение работы GPCR приводит к возникновению множества различных заболеваний, а сами рецепторы являются мишенью до 40 % выпускаемых лекарств. Точный размер надсемейства GPCR не известен, но почти 800 различных человеческих генов были предсказаны из анализа последовательности генома. Несмотря на многочисленные схемы, было предложено разделить надсемейство на три основных класса.

NMDA-рецептор — ионотропный рецептор глутамата, селективно связывающий N-метил-D-аспартат (NMDA).

Антагонисты NMDA рецептора, или NMDA-антагонисты — класс анестетиков, ингибирующих действие N-метил-D-аспартатного (NMDA) рецептора. NMDA-антагонисты часто применяются для анестезии животных, реже — человека, у которого они вызывают состояние так называемой диссоциативной анестезии. Исследования на грызунах показывают, что NMDA-антагонисты при чрезмерном использовании могут вызывать специфическое повреждение мозга — так называемые «лезии Олни», однако пока нет опубликованных данных о выраженности этой патологии у приматов.

В биохимии и фармакологии лиганд — это химическое соединение, которое образует комплекс с той или иной биомолекулой и производит, вследствие такого связывания, те или иные биохимические, физиологические или фармакологические эффекты. В случае связывания лиганда с белком лиганд обычно является малой сигнальной молекулой, связывающейся со специфическим участком связывания на белке-мишени. В случае связывания лиганда с ДНК лиганд обычно также является малой молекулой или ионом, или белком который связывается с двойной спиралью ДНК.

ГАМКА-рецептор — лиганд-зависимый ионный канал в химических синапсах нервной системы, который тормозит передачу нервного возбуждения и управляется с помощью ГАМК. Это, наряду с ГАМКС-рецептором, один из двух ионотропных ГАМК-рецепторов, отвечающий за реакцию организма на гамма-аминомасляную кислоту. Помимо места, связывающего ГАМК, рецепторный комплекс содержит аллостерические сегменты, способные связывать бензодиазепины, барбитураты, этанол, фуросемид, нейростероиды и пикротоксин.
5-HT2A-рецептор у млекопитающих является одним из подтипов 5-HT2-рецепторов, подсемейства серотониновых рецепторов. Он является метаботропным G-белок-сопряжённым рецептором. Рецептор этого подтипа (5-HT2A) является основным возбуждающим подтипом рецепторов среди всех G-белок-сопряжённых подтипов рецепторов для серотонина (5-HT). Однако рецепторы подтипа 5-HT2A способны, наоборот, оказывать ингибирующее, тормозящее воздействие в некоторых областях мозга, таких, как зрительная кора и орбитофронтальная кора.

Никотиновый ацетилхолиновый рецептор (н-холинорецептор, nACh-receptor) — подвид ацетилхолиновых рецепторов, который обеспечивает передачу нервного импульса через синапсы и активируется ацетилхолином, а также никотином. Этот рецептор вместе с ГАМКA-, ГАМКC- глициновым и 5-HT3-рецепторами образует семейство лиганд-зависимых ионных каналов с цистеиновой петлёй.
5-HT1A-рецептор — подтип серотониновых рецепторов подсемейства 5-HT1-рецепторов. Эндогенным лигандом-агонистом для них является моноаминовый нейромедиатор серотонин (5-гидрокситриптамин, 5-HT). Рецепторы этого подтипа относятся к семейству трансмембранных метаботропных G-белок-связанных рецепторов и связаны с гетеротримерным ингибиторным G-белком, так называемым Gi / Go. Этот подтип рецепторов опосредует тормозящую нейротрансмиссию. Ген, кодирующий белок этого рецептора у человека, обозначается HTR1A.

Внутренняя агонистическая активность или «внутренняя активность», «агонистическая активность», или «рецепторная эффективность», «эффективность по отношению к рецептору», коротко называемая «эффективность» в контексте фармакологии — термин, которым обозначают меру относительной способности комплекса лиганда с рецептором производить максимальный физиологический ответ.

Обра́тный агони́ст — это химическое соединение, которое связывается с тем же самым клеточным рецептором, что и агонист, однако производит физиологические эффекты, в целом противоположные физиологическим эффектам агониста.

В фармакологии термин части́чные агони́сты применяется по отношению к лекарствам и химическим соединениям, которые являются лигандами для конкретного подтипа клеточных рецепторов и способны активировать рецептор, то есть переводить его в активную пространственную конфигурацию, но с меньшей вероятностью, чем эндогенный агонист тех же рецепторов, рецепторная эффективность которого принимается за 100 % и который рассматривается, таким образом, как истинный полный агонист. Другими словами, внутренняя агонистическая активность частичного агониста по определению всегда больше 0 %, но меньше 100 %.
Конкурентные антагонисты — подтип лигандов-антагонистов рецепторов, которые обратимо связываются с рецепторами в том же самом участке связывания, что и физиологический эндогенный лиганд-агонист этого рецептора, но не вызывают активации рецептора. Физиологические агонисты и конкурентные антагонисты в этом случае «конкурируют» за связывание с одним и тем же участком связывания рецепторов. После того, как конкурентный антагонист свяжется с участком связывания рецептора, он предотвращает связывание с ним же агониста. Однако конкурентный антагонист не может ни «вытеснить» уже связавшийся с рецептором агонист из связи, ни предотвратить воздействие уже связавшегося агониста на клетку. Конечный результат конкуренции агонистов и антагонистов — и таким образом конечный уровень активности рецепторной системы — определяется соотношением молярных концентраций, относительным сродством к рецепторам и соотношением относительной внутренней агонистической активности агонистов и антагонистов. Поскольку высокие концентрации конкурентного антагониста повышают процент занятости рецепторов этим антагонистом, для достижения того же самого процента занятости рецепторов агонистом в этих условиях — и получения того же самого физиологического ответа — потребуются более высокие концентрации агониста, и наоборот — при более высоких концентрациях агониста требуется больше конкурентного антагониста для функциональной «блокады» рецепторов. В функциональных исследованиях конкурентные антагонисты вызывают параллельный сдвиг кривой «доза агониста-эффект» вправо, без изменения максимальной величины физиологического ответа.
Термин «неконкурентный антагонизм» используется для описания двух разных феноменов: в одном случае неконкурентный антагонист связывается с ортостерическим участком связывания лигандов на рецепторе, а в другом случае он связывается с аллостерическим участком связывания лигандов на рецепторе. И хотя механизм антагонистического действия различен в обоих случаях, они оба называются «неконкурентным антагонизмом», поскольку конечный результат воздействия антагониста в обоих случаях функционально очень похож. В отличие от конкурентных антагонистов, которые конкурируют с агонистами за занятость рецепторов и сдвигают кривую доза-эффект вправо, влияя на количество агониста, необходимое для получения максимального физиологического ответа, но никак не влияют на саму величину максимального физиологического ответа, неконкурентные антагонисты уменьшают величину максимального физиологического ответа, который может быть получен при любом сколь угодно большом количестве агониста. Это свойство и даёт им название «неконкурентные антагонисты», поскольку их эффект не может быть «уничтожен», обнулен или скомпенсирован увеличением количества агониста, сколь бы велико это увеличение ни было. В биологических системах, предназначенных для изучения влияния тех или иных антагонистов на рецепторы, неконкурентные антагонисты вызывают уменьшение «плато», и, в некоторых случаях, также сдвиг кривой вправо. Сдвиг кривой вправо происходит вследствие наличия во многих биологических рецепторных системах так называемого «рецепторного резерва», и ингибирование агонистического ответа под влиянием неконкурентного антагониста происходит только тогда, когда истощится (израсходуется) этот рецепторный резерв.
В фармакологии, физиологии и биохимии, термин эндогенный агонист по отношению к определённому подтипу клеточных рецепторов обозначает химическое соединение, которое отвечает следующим трём условиям:
- производится в самом организме или ткани в нормальных физиологических условиях, то есть является эндогенным веществом — например, нейромедиатором, цитокином или гормоном;
- является лигандом для данного подтипа рецепторов, то есть обладает способностью связываться с ортостерическим сайтом связывания данного подтипа рецепторов;
- обладает способностью после связывания активировать этот рецептор, изменяя его пространственную конфигурацию, что приводит к вызыванию соответствующего биохимического или физиологического ответа.

Лиганд-зависимые ионные каналы, лиганд-управляемые или лиганд-активируемые ионные каналы — также обычно называемые ионотропными рецепторами, представляют собой группу трансмембранных белков ионных каналов, которые позволяют ионам, например, Na+, K+, Ca2+ и/или Cl-, проходить через биологическую мембрану, посредством изменения конформации (открытия) в ответ на связывание химического мессенджера (то есть лиганда), такого как, например, молекулы нейромедиатора.
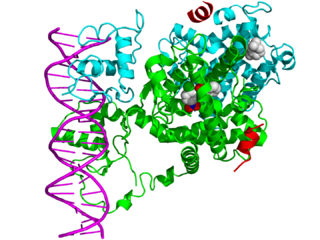
Ядерные рецепторы — это класс внутриклеточных белков, которые отвечают за восприятие стероидных и тиреоидных гормонов, а также некоторых других молекул. Уникальным свойством ядерных рецепторов, которое отличает их от других классов рецепторов, является их способность напрямую взаимодействовать с геномной ДНК и регулировать экспрессию соседних генов, тем самым контролируя развитие, гомеостаз и метаболизм организма. Следовательно, эти рецепторы классифицируются как факторы транскрипции. Регуляция экспрессии генов ядерными рецепторами обычно происходит только тогда, когда присутствует лиганд-молекула, которая влияет на поведение рецептора. Связывание лиганда с ядерным рецептором приводит к конформационному изменению рецептора, которое, в свою очередь, активирует рецептор, что приводит к усилению или понижению регуляции экспрессии гена.

В фармакологии и биохимии аллостерические модуляторы представляют собой группу веществ, которые связываются с рецептором, чтобы изменить реакцию этого рецептора на стимул. Некоторые из них, такие как бензодиазепины, являются наркотиками. Сайт, с которым связывается аллостерический модулятор, отличается от того, с которым связывался бы эндогенный агонист рецептора. Модуляторы и агонисты могут быть названы лигандами рецепторов.