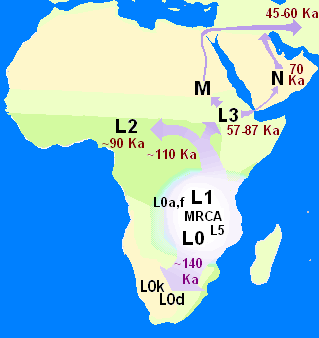
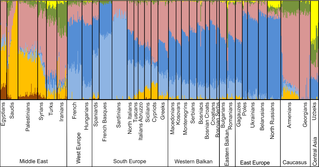
Генофонд казахов
Генофонд казахов — статистическая совокупность генных вариаций (гаплогрупп или совокупностей аллелей), присущая современным казахам.
Факты
Сведения о генофонде казахов противоречивы, что, возможно, обуславливается недостаточным количеством собранного генетического материала для достоверного обобщения. Согласно исследованиям двух разных групп исследователей (L. Castri & oth.[1] и D. Comas & oth.[2] цит. по[3]), 31,7—45 % митохондриальных гаплотипов казахского этноса имеют «западноевразийское» происхождение, 50—63,4 % имеют «восточноевразийское» происхождение, и 0—4,9 % характерны для населения Индии. Заметим, что в работе D. Comas & oth. были исследованы гаплотипы 232 индивидуумов из 12 популяций, среди прочих лишь 20 казахов, что является непредставительной выборкой для каких-либо выводов.
В работе Г. М. Березиной и др.[4] была собрана и исследована более представительная выборка из митохондриальных гаплотипов 246 казахов. По результатам, 58 % мтДНК казахов составили гаплогруппы D (17,9 %), C (16 %), G (16 %), A (3,25 %), F (2,44 %), предположительно восточноевразийского происхождения, и 41,46 % составили гаплогруппы H (13 %), T (4,07 %), J (4,07 %), K (4,07 %), U5 (3,25 %), I (0,41 %), V (0,81 %), W (1,63 %), предположительно западноевразийского происхождения.
В исследовании C. Lalueza-Fox et al.[3] были изучены митохондриальные гаплотипы 36 человеческих зубов из 15 могил, расположенных на территории Казахстана, возраст которых, определенный радиоуглеродным методом, варьирует от XIV—XI века до н. э. до III—V века н. э. Из этого числа только из 29 зубов удалось выделить ДНК, но и из них для 2 зубов препараты были забракованы. В 21 зубе были обнаружены европеоидные (западноевразийские) гаплотипы, а в 6 зубах — восточноевразийские. Отмечается динамика изменения гаплогрупп древнего населения Казахстана в сторону «востока Евразии». Так, согласно исследованиям, в XIV—X веке до н. э. во всех 8 зубах гаплогруппы западноевразийские, для периода VIII—VII веков до н. э. все 5 зубов также западноевразийские, для периода VII—III века до н. э. уже только 7 из 13 зубов относятся к западноевразийским. Единственный зуб относящийся к III—V векам н. э. оказался также западноевразийским.
На первый взгляд предыдущему исследованию противоречит работа R. Spencer Wells et al.[5], в которой собраны сведения по Y-хромосомным гаплотипам 54 этнических казахов и производится сравнительный анализ с типичными гаплотипами 49 этносов Евразии. Согласно результатам, наиболее распространённой мутацией гаплотипов казахов является M217 (гаплогруппа C (Y-ДНК)), характерная для восточно-азиатских этносов, например, японцев. Является загадкой, что самая распространённая мутация гаплотипов Евразии — M89 (гаплогруппа F (Y-ДНК)), не встречается лишь у казахов и японцев. Ещё более поразительно сильное отличие маркеров казахов и киргизов, традиционно считающихся близкими этносами. Наиболее распространенная мутация гаплотипов киргизов — M17 (гаплогруппа R1a (Y-ДНК)) более всего распространена среди славян, прибалтов, северных индийцев, афганцев и некоторых среднеазиатских народов. Все это вместе позволило авторам, по результатам кластеризации, заключить казахов в одну группу с азиатскими народами, причем на генетическом древе наиболее близким народом являются монголы[5].
Последние цифры сильно противоречат данным казахского ДНК-проекта[6]. В рамках данного проекта собраны Y-хромосомные гаплотипы 476 казахов. По этим данным, самые распространенные гаплогруппы: C (39 %), J (18 %), R (11 %), N (9 %), G (7 %), O (6 %). Также встречаются гаплогруппы Q (3 %), E (2 %), D (2 %), I (2 %), L (1 %). Таким образом, это явно противоречит предыдущим данным, а именно отсутствию у казахов гаплотипов макрогруппы F.
Однако и данные ДНК-проекта противоречат предыдущим результатам Э. К. Хуснутдиновой и др.[7]. Согласно им, распределение гаплогрупп Y-ДНК по выборке из 331 человека таково: C (25,3 %), J (18,2 %), N (15,2 %), R (10,1 %). К сожалению, данные про другим гаплогруппам не приведены.
По данным О. А. Балаганской, Y-хромосомная гаплогруппа С-M130 достигает у казахов 34,1 %, G-M201 — 23,7 %, O-M175 — 21,7 %, R1a1a-M198 — 6,8 %, N1c1-M178 — 3,5 %, J-M304 — 2,8 %, N1b — 1,2 %[8].
По данным крупнейшего исследования по казахскому геному Э. Хусаиновой и др.[9], изучавшего образцы 1171 представителя трех жузов, включая рода Торе и Кожа, наиболее распространенные Y-хромосомные гаплогруппы среди казахов: C2 (51,9%), R (12,8%), O2-M122 (8,03%), G (7,9%), N (6,9%), J (6,2%), Q (3,1%). Частота наиболее встречающейся гаплогруппы C2 варьируется от 11% (Канглы, Аргын) до 100% (Шакшам), при этом авторы отметили наличие не менее трех субкладов гаплогруппы С-M217: С-M401, C-M86, характерные для казахов Старшего, Младшего жуза и С-М407 рода Конырат (86%)в составе Среднего жуза соответственно. Вторая по распространенности гаплогруппа R имеет три субклада: R1a1a-M17 (6,5%), R1b-M343 (5,6%) и R2-M479 (0,8%). R1a1a-M17 наиболее характерна представителям родов Кожа (31,3%) и Ошакты (31%), R1b-M343 - представителям рода Кыпшак (36,8%) и R2-M479 - представителям родов Кожа (6,3%) и Жалайыр (5,4%). Гаплогруппа O2-M122 наиболее характерна для рода Найман (52,3%). Гаплогруппа G представлена субкладами G1-M285 (7,1%) и G2a-P15 (0,8%). G1-M285 наиболее характерна для рода Аргын (54%) и G2a-P15 - для рода Уак (18,2%). Гаплогруппа N-M231 часто встречается среди представителей родов Сиргели (64,1%), Уак (45,5%), Торе (16,7%) и Жалайыр (15,1%). Гаплогруппа J представлена субкладами J1-M267 и J2-M172. J1-M267 наиболее характерна для рода Ысты (39,4%) и J2-M172 - для рода Кожа (18,8%). Гаплогруппа Q чаще встречается среди представителей рода Канглы (66,7%).
По родам
Частота гаплогрупп в процентах среди казахских родов. Указаны больше 3 % в одном роду. Жирным выделены больше 40 %.[10]
| Гаплогруппа | Маркер | Казахи | Старший жуз | Средний жуз | Младший жуз | Аксуек | ||||||||||
| Уйсун | Жалайыр | Канлы | Аргын (включая таракты) | Кереи | Коныраты | Кыпшак | Найман | Уак | Алимулы | Байулы | Жетиру (союз семи родов) | Торе | Кожа- Сунак | |||
| количество выборок | N=1982 | N=248 | N=103 | N=27 | N=384 | N=102 | N=90 | N=133 | N=336 | N=45 | N=145 | N=130 | N=55 | N=28 | N=107 | |
| D* | M174 | 1 | 7 | |||||||||||||
| E1b1b1* | M35 | 4 | ||||||||||||||
| C2* | M217 | 17 | 50 | 38 | 7 | 3 | 66 | 10 | 13 | 25 | 36 | 7 | ||||
| C2b1a2 | M48 | 19 | 11 | 5 | 9 | 27 | 7 | 77 | 69 | 27 | 4 | 3 | ||||
| C2c1a1a1 | M407 | 4 | 86 | 4 | ||||||||||||
| G1 | M285 | 15 | 3 | 7 | 67 | 4 | 5 | 4 | 4 | 7 | ||||||
| G2a* | P15 | 4 | ||||||||||||||
| G2a2b2a | P303 | 4 | ||||||||||||||
| J1* | M267 | 3 | 12 | 4 | 4 | 4 | 7 | |||||||||
| J2* | M172 | 5 | 8 | 3 | 7 | 3 | 22 | 13 | 10 | |||||||
| J2a1b1 | M92 | 7 | ||||||||||||||
| N* | M231 | 3 | ||||||||||||||
| N1a1a | M178 | 5 | 4 | 22 | 3 | 64 | 3 | 7 | ||||||||
| O2* | M122 | 3 | ||||||||||||||
| O2a2b1 | M134 | 8 | 5 | 42 | 5 | 4 | ||||||||||
| Q* | M242 | 6 | 48 | 3 | 6 | 4 | ||||||||||
| Q1a2 | M143 | 5 | ||||||||||||||
| R1a1a* | M198 | 6 | 4 | 8 | 4 | 6 | 7 | 4 | 8 | 3 | 3 | 18 | 32 | |||
| R1a1a1b1a1 | M458 | 3 | ||||||||||||||
| R1b1a1a* | P297 | 4 | 4 | 47 | 4 | |||||||||||
| R1b1a1a2 | M269 | 7 | 5 | 4 | ||||||||||||
| R2a | M124 | 25 | 10 | |||||||||||||
| L1a2 | M357 | 4 | ||||||||||||||
| L1b | M317 | 4 | ||||||||||||||
| Разнообразие (GD) | 89 | 72 | 80 | 76 | 54 | 56 | 27 | 72 | 74 | 58 | 40 | 50 | 85 | 80 | 87 | |
Гаплогруппы Старшего жуза
Старший Жуз сформирован сочетанием не только генетически родственных племенных групп, но и генетически удалённых[11].
Y гаплогруппы родов Старшего жуза в процентах[11].
| № | Гаплогруппа | Старший жуз | Жалайыр | Дулаты | Албаны | Суаны | Сарыуйсуны | Ошакты | Шапрашты | Шанышкылы | Шакшам | Канлы | Сиргели | Ысты |
| 1 | C3(x) | 2,1 | 1,1 | 3,1 | 2,2 | 0,0 | 0,0 | 0,0 | 6,7 | 0,0 | 0,0 | 7,5 | 0,0 | 0,0 |
| 2 | C3-M401 | 45,0 | 41,3 | 62,3 | 65,2 | 61,0 | 87,5 | 36,7 | 46,7 | 53,8 | 100,0 | 5,0 | 15,6 | 3,5 |
| 3 | C3-M407 | 0,2 | 0,0 | 0,0 | 2,2 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| 4 | C3c-M86 | 5,3 | 4,3 | 8,9 | 8,7 | 0,0 | 0,0 | 0,0 | 6,7 | 15,4 | 0,0 | 2,5 | 0,0 | 1,8 |
| 5 | E1b-M35 | 0,7 | 1,1 | 0,5 | 0,0 | 0,0 | 0,0 | 3,3 | 0,0 | 0,0 | 0,0 | 0,0 | 3,1 | 0,0 |
| 6 | G(x) | 0,2 | 0,0 | 0,0 | 2,2 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| 7 | G1-M285 | 2,3 | 5,4 | 1,0 | 2,2 | 0,0 | 0,0 | 10,0 | 0,0 | 0,0 | 0,0 | 2,5 | 0,0 | 1,8 |
| 8 | G2-P287 | 1,6 | 6,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 6,7 | 0,0 | 0,0 | 2,5 | 0,0 | 1,8 |
| 9 | I1-M253• | 0,4 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 3,5 |
| 10 | I2a-L460• | 1,2 | 2,2 | 1,0 | 0,0 | 0,0 | 0,0 | 6,7 | 6,7 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| 11 | I2b-L415• | 0,7 | 0,0 | 2,1 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| 12 | J(x) | 0,7 | 0,0 | 0,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 15,4 | 0,0 | 2,5 | 0,0 | 0,0 |
| 13 | J1-M267• | 7,9 | 3,3 | 1,6 | 0,0 | 0,0 | 12,5 | 0,0 | 13,3 | 0,0 | 0,0 | 0,0 | 0,0 | 63,2 |
| 14 | J2a-M410• | 5,6 | 14,1 | 7,9 | 0,0 | 0,0 | 0,0 | 0,0 | 6,7 | 0,0 | 0,0 | 2,5 | 3,1 | 1,8 |
| 15 | L• | 0,4 | 0,0 | 0,0 | 0,0 | 4,9 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| 16 | N(x) | 0,2 | 1,1 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| 17 | N-P43 | 0,4 | 0,0 | 0,5 | 0,0 | 0,0 | 0,0 | 0,0 | 6,7 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| 18 | N-M46 | 6,3 | 9,8 | 1,0 | 4,3 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 65,6 | 3,5 |
| 19 | O(xM122) | 1,1 | 0,0 | 0,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 8,8 |
| 20 | O-M122(xM134) | 0,2 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 3,1 | 0,0 |
| 21 | O-M134 | 1,1 | 1,1 | 1,0 | 2,2 | 2,4 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 3,1 | 0,0 |
| 22 | Q | 5,3 | 0,0 | 0,5 | 2,2 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 67,5 | 3,1 | 0,0 |
| 23 | R(x) | 0,4 | 0,0 | 0,5 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 2,5 | 0,0 | 0,0 |
| 24 | R1a1a-M198 | 6,5 | 3,3 | 3,7 | 6,5 | 31,7 | 0,0 | 26,7 | 0,0 | 7,7 | 0,0 | 5,0 | 0,0 | 0,0 |
| 25 | R1b-M478 | 2,1 | 0,0 | 2,1 | 0,0 | 0,0 | 0,0 | 6,7 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 10,5 |
| 26 | R1b-M269 | 0,5 | 0,0 | 0,0 | 2,2 | 0,0 | 0,0 | 3,3 | 0,0 | 0,0 | 0,0 | 0,0 | 3,1 | 0,0 |
| 27 | R2-M124* | 1,4 | 5,4 | 0,5 | 0,0 | 0,0 | 0,0 | 3,3 | 0,0 | 7,7 | 0,0 | 0,0 | 0,0 | 0,0 |
| 28 | не определено | 0,4 | 0,0 | 0,5 | 0,0 | 0,0 | 0,0 | 3,3 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 | 0,0 |
| число пробантов | 567 | 92 | 191 | 46 | 41 | 8 | 30 | 15 | 13 | 2 | 40 | 32 | 57 | |
| Генетическое разнообразие | 0.773 ±0,017 | 0.793 ±0,036 | 0.597 ±0,041 | 0.570 ±0,085 | 0.538 ±0,059 | 0.250 ±0,18 | 0.798 ±0,052 | 0.791 ±0,105 | 0.705 ±0,122 | 0 | 0.544 ±0,094 | 0.557 ±0,097 | 0.588 ±0,073 |
См. также
Примечания
- ↑ L. Castri, P. Francalacci, L. Simoni, D. Luiselli, F. Facchini and D. Pettener, unpublished data
- ↑ http://www.upf.edu/bioevo/2004Jaume/JB2004-Comas-EJHG.pdf (недоступная ссылка)
- ↑ 1 2 http://www.upf.edu/bioevo/2004BioEvo/BE2004-Lalueza-Fox-RSL.pdf (недоступная ссылка)
- ↑ Полиморфизм митохондриальной ДНК в казахской популяции. Дата обращения: 7 сентября 2010. Архивировано 21 июня 2020 года.
- ↑ 1 2 Архивированная копия. Дата обращения: 7 ноября 2009. Архивировано 5 октября 2014 года.
- ↑ База Данных ДНК-паспортов казахов — Шежире / Билингво / ДНК-проект. Дата обращения: 7 сентября 2010. Архивировано из оригинала 1 февраля 2015 года.
- ↑ 10_1. Дата обращения: 8 сентября 2010. Архивировано 19 августа 2019 года.
- ↑ Балаганская О. А. Полиморфизм Y-хромосомы у тюркоязычного населения Алтая, Саян, Тянь-Шаня и Памира в контексте взаимодействия генофондов Западной и Восточной Евразии // Учреждение Российской академии медицинских наук «Медико-генетический научный центр РАМН». — 2011. (Таблица 2. Частоты гаплогрупп Y хромосомы у изученных крупных народов, этно-территориальных объединений и регионов Евразии)
- ↑ Elmira Khussainova, Ilya Kisselev, Olzhas Iksan, Bakhytzhan Bekmanov, Liliya Skvortsova. Genetic Relationship Among the Kazakh People Based on Y-STR Markers Reveals Evidence of Genetic Variation Among Tribes and Zhuz // Frontiers in Genetics. — 2022. — Т. 12. — ISSN 1664-8021. — doi:10.3389/fgene.2021.801295/full.
- ↑ Архивированная копия. Дата обращения: 19 февраля 2018. Архивировано 9 июля 2021 года.
- ↑ 1 2 Molecular Genetic Analysis of Population Structure of the Great Zhuz Kazakh Tribal Union Based on Y-Chromosome Polymorphism | SpringerLink. Дата обращения: 26 ноября 2018. Архивировано 9 июля 2021 года.
Ссылки
- АНАЛИЗ ЖУЗОВОЙ ОРГАНИЗАЦИИ КАЗАХСКИХ ПЛЕМЕН НА ОСНОВЕ РАСПРЕДЕЛЕНИЯ ГАПЛОГРУПП Y-ХРОМОСОМЫ
- Генетика русских украинцев белорусов и татар, славян и кавказцев, евреев, финнов и других популяций.
| Старший жуз | |
|---|---|
| Средний жуз | |
| Младший жуз | |
| Аксуйек | |
| Не входят в жузы | |
| Другое | |
| Ключевые понятия |  | |
|---|---|---|
| Области генетики | ||
| Закономерности | ||
| Связанные темы | ||