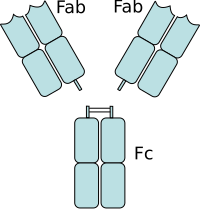
Антитела́, иммуноглобулины — крупные глобулярные белки плазмы крови, выделяющиеся плазматическими клетками иммунной системы и служащие для нейтрализации клеток патогенов и вирусов, а также белковых ядов и некоторых других чужеродных веществ. Каждое антитело распознаёт уникальный элемент патогена, отсутствующий в самом организме, — антиген, а в пределах данного антигена — определённый его участок, эпитоп. Связываясь с антигенами на поверхности патогенов, антитела могут либо непосредственно нейтрализовать их, либо привлекать другие компоненты иммунной системы, такие как система комплемента и фагоциты, чтобы уничтожить чужеродные клетки или вирусные частицы. Антитела — важнейший компонент гуморального специфического иммунитета.

Систе́ма комплеме́нта — комплекс защитных белков, постоянно присутствующих в крови. Это каскадная система протеолитических ферментов, предназначенная для гуморальной защиты организма от действия чужеродных агентов, она участвует в реализации иммунного ответа организма. Является важным компонентом как врождённого, так и приобретённого иммунитета. Выделяют три основных пути активации системы комплемента: классический, альтернативный и лектиновый. Для запуска классического пути комплемента необходима опсонизация чужеродной клетки антителами, а альтернативный и лектиновый пути могут активироваться в отсутствие антител. Поздние стадии у всех трёх путей активации системы комплемента одинаковы и включают образование мембраноатакующего комплекса, который нарушает целостность мембраны клетки-патогена и приводит к её гибели.
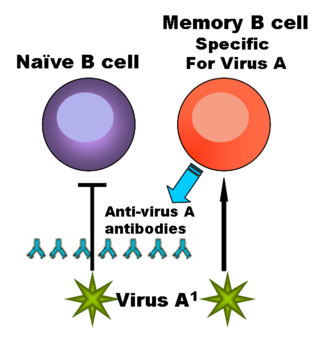
— функциональный тип лимфоцитов, играющих важную роль в обеспечении гуморального иммунитета. При контакте с антигеном или стимуляции со стороны T-клеток некоторые B-лимфоциты трансформируются в плазматические клетки, способные к продукции антител. Другие активированные B-лимфоциты превращаются в B-клетки памяти. Помимо продукции антител, В-клетки выполняют множество других функций: выступают в качестве антигенпрезентирующих клеток, продуцируют цитокины и экзосомы.
Приобретённый иммунитет — способность организма обезвреживать чужеродные и потенциально опасные микроорганизмы, которые уже попадали в организм ранее. Представляет собой результат работы системы высокоспециализированных клеток (лимфоцитов), расположенных по всему организму. Считается, что система приобретённого иммунитета возникла у челюстноротых позвоночных. Она тесно взаимосвязана с гораздо более древней системой врождённого иммунитета, которая является основным средством защиты от патогенных микроорганизмов у большинства живых существ.

Лёгкие цепи иммуноглобулинов — это белковые субъединицы иммуноглобулинов. Молекулы иммуноглобулинов класса G состоят из двух лёгких и двух тяжёлых цепей. Лёгкие цепи иммуноглобулинов имеют молекулярную массу около 28 кДа.

Участок связывания антигена — участок молекулы иммуноглобулина, который связывает антиген. Он состоит из одного константного и одного вариабельного домена легкой и тяжелой цепей. Эти домены образуют паратоп — сайт связывания антигена на N-конце. Вариабельные домены связывают эпитоп специфических антигенов.
Бело́к A — белок молекулярной массой 40—60 кДа, выделенный с поверхности клеточной стенки золотистого стафилококка. Белок A используется в биохимических исследованиях, так как хорошо связывает многие иммуноглобулины млекопитающих, особенно иммуноглобулины G.
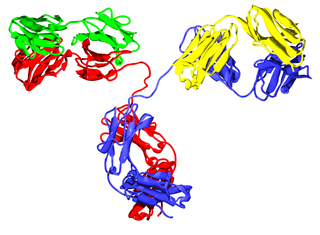
Тяжёлые цепи иммуноглобулинов — это большие полипептидные субъединицы иммуноглобулинов (антител).

Гиперчувствительность — повышенная чувствительность организма к какому-либо веществу. Гиперчувствительность является нежелательной излишней реакцией иммунной системы и может привести не только к дискомфорту, но и к смерти.

V(D)J-рекомбина́ция, или V(D)J-реаранжиро́вка , — механизм соматической рекомбинации ДНК, происходящий на ранних этапах дифференцировки лимфоцитов и приводящий к формированию антиген-распознающих участков антител и Т-клеточного рецептора. Гены иммуноглобулинов и Т-клеточных рецепторов состоят из повторяющихся сегментов, принадлежащих к трём классам: V, D и J. В процессе V(D)J-перестройки генные сегменты, по одному из каждого класса, соединяются вместе. Объединённая последовательность сегментов V(D)J кодирует вариабельные домены каждой из цепей рецептора или антитела.
CD64 — мембранный белок, гликопротеин, Fc-рецептор к мономерным иммуноглобулинам изотипа IgG с высокой аффинностью. Обозначается как Fc-гамма-рецептор 1 (FcγRI). После связывания IgG белок CD64 взаимодействует с гамма-цепью (γ-цепью), в которой расположен мотив ITAM, запускающий клеточную активацию. Существуют 3 типа CD64: CD64A, CD64B и CD64C.
Неонатальный Fc-рецептор — рецептор Fc-фрагмента IgG, продукт гена человека FCGRT. Это гетеродимерный рецептор, состоящий из тяжёлой цепи с MHC I-подобной укладкой и микроглобулина β2. Рецептор узнаёт участок, соединяющий CH2- и CH3-домены IgG, что отличает его от классического Fc-рецептора γ и компонента системы комплемента C1q и сближает с бактериальными белками A и G, которые связываются с тем же участком. На ранних этапах жизни организма неонатальный Fc-рецептор обеспечивает усвоение антител IgG из крови или молока матери, а во взрослом организме играет важную роль в поддержании гомеостаза антител этого изотипа и сывороточного альбумина, защищая их от преждевременной деградации. У людей данный рецептор обнаруживается в эндотелии сосудов, профессиональных антигенпрезентирующих клетках, эпителии кишечника, эндотелии центральной нервной системы, подоцитах почек и в лёгких.

Иммуноглобули́ны G (IgG) — класс антител. IgG составляют около 75 % антител плазмы крови у человека и являются наиболее часто встречаемыми в кровотоке антителами. IgG продуцируются плазматическими B-клетками, и каждая молекула IgG имеет два сайта связывания антигена.

Иммуноглобули́ны M (IgM) — класс антител. Молекулы IgM являются самыми тяжёлыми и наиболее сложно организованными иммуноглобулинами. Молекула свободного IgM представляет собой пентамер, каждый мономер которого состоит из двух тяжёлых цепей (μ-цепей) и двух лёгких цепей κ- или λ-типов. Мономеры объединены в пентамер посредством дисульфидных мостиков и J-цепи. При первичном попадании в организм антигена иммуноглобулины IgM из всех антител образуются первыми. Кроме того, они первыми появляются в онто- и филогенезе. У человека и других млекопитающих IgM синтезируются плазматическими клетками, находящимися в селезёнке. IgM наиболее активны в антибактериальном иммунитете и при ряде аутоиммунных заболеваний.

Иммуноглобули́ны A (IgA) — класс антител. IgA доминируют в секретах организма, их доля в плазме крови составляет 10—15 % от общего количества всех иммуноглобулинов. За сутки в просвет кишечника у человека выделяется от 3 до 5 г IgA. У человека имеются два подкласса IgA: IgA1 и IgA2. IgA присутствуют в организме преимущественно в мономерной и димерной форме. Молекулы IgA1 наиболее многочисленны в плазме крови, а IgA2 — в секретах. Соотношение клеток, секретирующих IgA1 и IgA2, различно в разных лимфоидных тканях.
Иммуноглобули́ны Y (IgY) — класс антител, имеющийся у птиц, рептилий и амфибий. IgY накапливаются в большом количестве в желтке яиц, куда транспортируются из плазмы крови. Молекула IgY состоит из двух тяжёлых и двух лёгких цепей. В ранних источниках IgY ошибочно именуют IgG из-за структурного и функционального сходства с иммуноглобулинами G млекопитающих. Однако между IgY и IgG всё же имеются существенные структурные и функциональные различия, а антитела, направленные против IgG (анти-IgG), не действуют на IgY.

Класси́ческий пу́ть актива́ции систе́мы комплеме́нта — один из трёх путей активации системы комплемента, наряду с альтернативным путём и лектиновым путём. Классический путь запускают комплексы антиген-антитело, причём антитела должны принадлежать к классу IgG или IgM. После активации происходит сборка белкового комплекса C3-конвертазы (C4b2b), который разрезает белок комплемента C3. Один из фрагментов C3, C3b, связывается с C3-конвертазой, в результате чего образуется C5-конвертаза (C4b2b3b). C5-конвертаза разрезает компонент комплемента 5, после чего образовавшиеся белковые фрагменты привлекают фагоциты в очаг инфекции и способствуют поглощению клеток патогенов. C5-конвертаза также запускает финальные стадии каскада комплемента, которые завершаются формированием мембраноатакующего комплекса. Он формирует поры в мембране клетки микроорганизма, вызывая её лизис и гибель. Помимо микробных клеток, классический путь комплемента могут запускать апоптотические тельца и некротизированные клетки.

Переключе́ние кла́ссов антите́л, или переключе́ние кла́ссов иммуноглобули́нов, или переключе́ние изоти́пов, — процесс переключения B-лимфоцита (B-клетки) с синтеза антител одного класса на синтез антител другого класса, например, с иммуноглобулинов M (IgM) на иммуноглобулины G (IgG). В ходе переключения классов константная часть локуса IGH, кодирующего тяжёлую цепь антител, претерпевает изменения, в то время как вариабельный участок остаётся неизменным. Так как вариабельный участок антитела не меняется, его антигенная специфичность остаётся прежней, и антитело продолжает распознавать тот же эпитоп.
Аллоти́п — вариация иммуноглобулина которая присутствует среди классов антител и проявляется гетерогенностью иммуноглобулинов, присутствующих у одного вида позвоночных. Структура полипептидной цепи иммуноглобулина диктуется и контролируется количеством генов, кодируемых в зародышевой линии. Однако эти гены, как было обнаружено серологическими и химическими методами, могут быть высоко полиморфными. Этот полиморфизм впоследствии проецируется на общую аминокислотную структуру цепей антител. Полиморфные эпитопы могут присутствовать в постоянных областях иммуноглобулина как на тяжелых, так и на легких цепях, различаясь между отдельными людьми или этническими группами и в некоторых случаях могут представлять собой иммуногенные детерминанты. Воздействие на людей несамостоятельного аллотипа может вызвать антиаллотипический ответ и стать причиной проблем, например, у пациента после переливания крови или у беременной женщины. Однако важно отметить, что не все вариации аминокислотной последовательности иммуноглобулина представляют собой детерминант, ответственный за иммунный ответ. Некоторые из этих аллотипических детерминант могут присутствовать в местах, которые не очень хорошо подвержены воздействию и поэтому вряд ли могут быть серологически различимы. В других случаях вариация одного изотипа может быть компенсирована присутствием этого детерминанта на другом изотипе антител у одного индивидуума. Это означает, что дивергентный аллотип тяжелой цепи IgG-антитела может быть сбалансирован присутствием этого аллотипа на тяжелой цепи, например, IgA-антитела и поэтому называется изоаллотипическим вариантом.особенно большое количество полиморфизмов было обнаружено в подклассах IgG-антител. Особенно большое количество полиморфизмов было обнаружено в подклассах IgG антител. Которые практически использовались в судебной медицине и в тестировании отцовства, прежде чем были заменены современной ДНК-дактилоскопией.
CD16 — мембранный белок суперсемейства иммуноглобулинов, низкоаффинный рецептор для Fc-фрагмента иммуноглобулинов G (IgG). Экспрессирован на поверхности естественных киллеров, нейтрофилов, моноцитов, макрофагов и определённых T-лимфоцитов. Существуют две формы CD16: FcγRIIIa (CD16a) и FcγRIIIb (CD16b), которые участвуют в переносе сигнала. Это наиболее изученный мембранный рецептор, играющий роль в антитело-зависимой клеточной цитотоксичности (ADCC).