Азотфикса́ция, или азотофиксация, — фиксация молекулярного атмосферного азота, диазотрофия. Процесс восстановления молекулы азота и включения её в состав своей биомассы прокариотными микроорганизмами. Важнейший источник азота в биологическом круговороте. В наземных экосистемах азотфиксаторы локализуются в основном в почве.
Денитрификация — сумма микробиологических процессов восстановления нитратов до нитритов и далее до газообразных оксидов и молекулярного азота. В результате их азот возвращается в атмосферу и становится недоступным большинству организмов. Осуществляется только прокариотами в анаэробных условиях и связана с получением ими энергии.

Актиномицеты — порядок бактерий, имеющих способность к формированию на некоторых стадиях развития ветвящегося мицелия, которая проявляется у них в оптимальных для существования условиях. Некоторые исследователи, подчёркивая бактериальную природу актиномицетов, называют их аналог грибного мицелия тонкими нитями; их диаметр 0,4—1,5 мкм. Актиномицеты имеют кислотоустойчивый тип клеточной стенки, которая окрашивается по Граму как грамположительно, однако по структуре ближе к грамотрицательным. Характеризуются высоким (60—75 %) содержанием ГЦ пар в ДНК.
Нитрификация — микробиологический процесс окисления аммиака до азотистой кислоты или её самой далее до азотной кислоты, что связано либо с получением энергии, либо с защитой от активных форм кислорода, образующихся при разложении пероксида водорода.

Анаэробы — организмы, получающие энергию при отсутствии доступа кислорода путём субстратного фосфорилирования, конечные продукты неполного окисления субстрата при этом могут быть окислены с получением большего количества энергии в виде АТФ.
Анаэробное дыхание — это биохимический процесс окисления органических субстратов или молекулярного водорода с использованием в дыхательной ЭТЦ в качестве конечного акцептора электронов вместо O2 других окислителей неорганической или органической природы. Как и в случае аэробного дыхания, выделяющаяся в ходе реакции свободная энергия запасается в виде трансмембранного протонного потенциала, использующегося АТФ-синтазой для синтеза АТФ.
Метаногенез, биосинтез метана — процесс образования метана анаэробными археями, сопряжённый с получением ими энергии. Существует три типа метаногенеза:
- Восстановление одноуглеродных соединений с помощью молекулярного водорода или двух- и более углеродных спиртов.
- Диспропорционирование одноуглеродных соединений.
- Диспропорционирование ацетата.
Анаэробное окисление метана — процесс окисления метана до углекислого газа, производимый некультивируемыми археями групп ANME-1, ANME-2 и ANME-3, близкими к Methanosarcinales при отсутствии в среде молекулярного кислорода. Биохимия и распространённость процесса в природе изучены пока недостаточно.

Сульфатредуцирующие бактерии представляют собой группу бактерий, характеризующуюся способностью окислять сероводород и отлагать в своём теле крупинки серы. Признак этой группы, как видно из сказанного, чисто физиологический, морфологических же признаков эта группа не имеет. Известны учёным серные бактерии уже давно. Ф. Кон первый изучал их физиологию, и так как серные бактерии всегда были находимы в тех местах, где наблюдалось выделение сероводорода, то Кон пришёл к заключению, что образование сероводорода находится в зависимости от жизнедеятельности этих бактерий, разрушающих различные соединения, заключающие в себе серу, и выделяющих сероводород. В этом своём заключении он опирается на наблюдения Лотара Мейера, который заметил, что вода Линдекских минеральных источников с серными бактериями — «водорослями», как он думал — после четырёхмесячного стояния заключает гораздо больше сероводорода, чем без «водорослей», и что «водоросли» восстанавливают, по-видимому, сернокислые соли в сероводород. Наблюдения над минеральными источниками в Иоганисбаде показали, далее, Кону, что там, где нет серных бактерий (Beggiatoa), там нет и сероводорода, и наоборот; кроме того, исследуя материал, присланный ему с берегов Дании Вормигом из мест, где замечалось выделение сероводорода, Кон нашёл много спирилл и монад с отложением серы внутри их тела, что ещё более укрепило его во взгляде на способность многих микроорганизмов выделять сероводород. Гоппе-Зейлер первый усомнился в справедливости заключений Кона, а Виноградский блестяще доказал ошибочность взглядов Кона; взгляд Виноградского считается теперь общепризнанным. По его мнению, серные бактерии не имеют никакого отношения к образованию сероводорода и восстановлению сульфатов, отложение же серы внутри тела бактерий Виноградский принимает как следствие окисления находящегося в воде сероводорода бактериями. Шаблон:Биофото Для получения серных бактерий в культурах Виноградский разрезал на мелкие куски свежевыкопанное корневище водяного растения — лучше всего для этой цели ему служило корневище сусака — и клал их в высокий сосуд, в который наливал колодезной воды с прибавкой небольшого количества гипса. Уже по прошествии 5—6 дней вода сосуда начинала пахнуть сероводородом, выделение его понемногу увеличивалось, но серных бактерий заметно ещё не было. Только через 4 недели можно, наконец, заметить появление нитей Beggiatoa, а месяца через 2 стенки сосуда вблизи поверхности воды оказывались уже вполне ими покрыты. Если кусочки Butomus umbellatus перед тем, как бросить в воду, подержать некоторое время в кипящей воде, то хотя сероводород и образуется потом в сосуде с гипсом, но Beggiatoa не появляется. Таким образом Виноградский мог заключить, что не Beggiatoa образует сероводород, но газ этот появляется помимо серных бактерий. Сера отлагается в теле бактерий только тогда, когда бактерии развиваются в воде, заключающей сероводород, и исчезает уже по прошествии 24 часов, когда бактерии переносят в ключевую или кипячёную воду. В случае недостатка сероводорода в воде бактерии окисляют находящуюся в их клетках серу до серной кислоты, которая образует с углекислой известью окружающей воды гипс. Схематически процессы, происходящие при этом, можно представить в виде следующих формул:

 .
.

Протеобактерии — наиболее многочисленная группа бактерий — 1534 вида или примерно треть от всех известных видов бактерий.
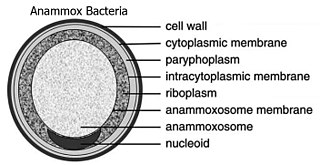
Candidatus Brocadia anammoxidans (лат.) — вид анаэробных хемолитоавтотрофных кокковидных бактерий рода Candidatus Brocadia, впервые выделен из сооружений по очистке сточных вод в Делфте, Нидерланды, также были обнаружены в Чёрном море.

Methylococcus capsulatus (лат.) — облигатно метанотрофный грамотрицательный кокк рода Methylococcus, использует метан в качестве единственного источника углерода, использование метана ингибируется присутствием ионов аммония.
Литотрофы — организмы, для которых донорами электронов, необходимых для многих клеточных процессов, являются неорганические вещества. Противопоставлены органотрофам.

Acetobacteraceae (лат., возможное русское название — уксуснокислые бактерии) — семейство бактерий из типа протеобактерий, которые получают энергию, окисляя этанол до уксусной кислоты (являются строго облигатными аэробами). Поэтому они часто развиваются вслед за дрожжами, используя продукт спиртового брожения как субстрат для роста. Это грамотрицательные аэробные палочковидные бактерии, слабоподвижные за счёт перитрихиально или полярно расположенных жгутиков или неподвижные. Довольно требовательны к субстратам для роста. Почти все виды нуждаются в отдельных витаминах, в первую очередь в пантотеновой кислоте, однако есть формы, способные к синтезу всех факторов роста. Не стоит путать их с родом Acetobacterium, которые являются анаэробными гомоацетогенными факультативными автотрофами (ацетогенами) и могут восстанавливать CO2 до уксусной кислоты (см. статью Acetobacterium woodii). Для производства уксуса применяются в основном представители двух родов — Gluconobacter и Acetobacter.

Метилотрофы, или Метилобактерии — аэробные или анаэробные бактерии, использующие в качестве источников углерода и энергии окисленные или замещённые производные метана, не имеющие С-С связи, но неспособные расти на самом метане. Ростовыми субстратами для метилобактерий служат метанол, метиламин, диметиламин, триметиламин, галометаны, серосодержащие соединения — метансульфоновая кислота, диметилсульфид и многие другие. Некоторые из этих соединений, например, триметиламин — (СН3)N, содержат более одного атома углерода, но не имеют С-С связи. Метилобактерии, в отличие от метанотрофов, не имеют сложной системы внутрицитоплазматических мембран (ВЦМ). По типу питания различают три группы метилобактерий:
- Облигатные — растут только на С1-соединениях;
- Ограниченно-факультативные — используют наряду с С1-субстратами одно или несколько полиуглеродных (Сn) соединений;
- Факультативные — используют, кроме С1-соединений, широкий спектр Сn-соединений.
Путь Э́нтнера — Ду́дорова, или КДФГ-путь — путь окисления глюкозы, приводящий к образованию из одной молекулы глюкозы двух молекул пирувата, одной молекулы АТФ и двух молекул восстановленных пиридиновых нуклеотидов. Хотя ранее считалось, что он имеет место лишь у небольшого числа грамотрицательных бактерий, в настоящее время установлено, что этот путь распространён в природе чрезвычайно широко и используется различными группами грамположительных и грамотрицательных бактерий, а также некоторыми археями и даже эукариотами.
Ацетогены — прокариоты, которые выделяют ацетат (CH3COO−) в качестве конечного продукта анаэробного дыхания или гомоацетатного брожения. Однако обычно этот термин используется в более узком смысле: только в отношении бактерий и архей, которые одновременно осуществляют анаэробное дыхание и фиксацию углерода в восстановительном ацетил-КоА-пути (путь Вуда — Льюнгдаля). Этот процесс известен как ацетогенез. Ацетогенов следует отличать от уксуснокислых бактерий, которые выделяют ацетат, получающийся в результате неполного аэробного окисления этилового спирта до уксусной кислоты.
Карбоксидобактерии — группа аэробных хемолитоавтотрофных бактерий, получающих энергию при окислении угарного газа (СО), объединяемых на основании физиологических особенностей. Не являются таксономической группой. Включают α, β, и γ-протеобактерии, фирмикутов, и актинобактерий. Большинство известных карбоксидобактерий являются грамотрицательными бактериями.
Говард Далтон (1944—2008) — британский биохимик и микробиолог.
Джон Родни Куэйл — британский биохимик и микробиолог.
