Метаболи́зм, или обме́н веще́ств, — это химические реакции, поддерживающие жизнь в живом организме. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды.
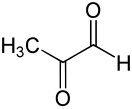
Глико́лиз, или путь Эмбдена — Мейергофа — Парнаса — процесс окисления глюкозы, при котором из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты. Гликолиз состоит из цепи последовательных ферментативных реакций и сопровождается запасанием энергии в форме АТФ и НАДH. Гликолиз является универсальным путём катаболизма глюкозы и одним из трёх путей окисления глюкозы, встречающихся в живых клетках. Реакция гликолиза в суммарном виде выглядит следующим образом:
- Глюкоза + 2НАД+ + 2АДФ + 2Pi → 2 пируват + 2НАД*H + 2Н+ + 2АТФ + 2Н2O.

Цитозо́ль — жидкое содержимое клетки. Большую часть цитозоля занимает внутриклеточная жидкость. Цитозоль разбивается на компартменты при помощи разнообразных мембран. У эукариот цитозоль располагается под плазматической мембраной и является частью цитоплазмы, в которую, помимо цитозоля, входят митохондрии, пластиды и другие органеллы, но не содержащаяся в них жидкость и внутренние структуры. Таким образом, цитозоль представляет собой жидкий матрикс, окружающий органеллы. У прокариот большая часть химических реакций метаболизма происходит в цитозоле, и лишь небольшая их часть происходит в мембранах и периплазматическом пространстве. У эукариот, хотя многие реакции протекают в органеллах, некоторые реакции, например, гликолиз или синтез жирных кислот, происходят в цитозоле.
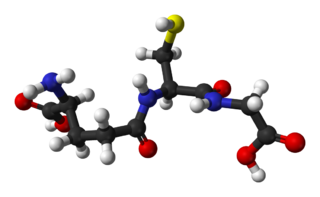
Глутатион — это трипептид γ-глутамилцистеинилглицин. Глутатион содержит необычную пептидную связь между аминогруппой цистеина и карбоксильной группой боковой цепи глутамата. Значение глутатиона в клетке определяется его антиоксидантными свойствами. Фактически глутатион не только защищает клетку от токсичных свободных радикалов, но и в целом определяет окислительно-восстановительные характеристики внутриклеточной среды.

Окисли́тельное фосфорили́рование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Хотя различные формы жизни на Земле используют разные питательные вещества, АТФ является универсальным соединением, в котором запасается энергия, необходимая для других метаболических процессов. Почти все аэробные организмы осуществляют окислительное фосфорилирование. Вероятно, широкому распространению этого метаболического пути способствовала его высокая энергетическая эффективность по сравнению с анаэробным брожением.

Убиквити́н — небольшой консервативный белок эукариот, участвующий в регуляции процессов внутриклеточной деградации других белков, а также в модификации их функций. Он присутствует почти во всех тканях многоклеточных эукариот, а также у одноклеточных эукариотических организмов. Убиквитин был открыт в 1975 году Гидеоном Голдштейном с соавторами и охарактеризован в 70—80-х годах XX века. В геноме человека есть четыре гена, кодирующих убиквитин: UBB, UBC, UBA52 и RPS27A.

Фосфоинозитид-3-киназы, или фосфатидилинозитол-3-киназы — семейство ферментов, фосфорилирующих фосфатидилинозитол в положении 3D инозитольного кольца. Являются ключевым элементом PI3K сигнального пути.

Тилакоиды — ограниченные мембраной компартменты внутри хлоропластов и цианобактерий. В тилакоидах происходят светозависимые реакции фотосинтеза. Слово «тилакоид» происходит от греческого слова θύλακος, означающего «мешочек». Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Тилакоиды хлоропластов часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами. Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.

Факторы транскрипции — белки́, контролирующие процесс синтеза мРНК, а также других видов РНК на матрице ДНК (транскрипцию) путём связывания со специфичными участками ДНК. Транскрипционные факторы выполняют свою функцию либо самостоятельно, либо в комплексе с другими белками. Они обеспечивают снижение (репрессоры) или повышение (активаторы) константы связывания РНК-полимеразы с регуляторными последовательностями регулируемого гена.
CD9 — мембранный белок, гликопротеин из надсемейства тетраспанинов. Белок был открыт в 1981 году и стал первым представителем семейства белков тетраспанинов, включающим на настоящий момент 34 белка млекопитающих, из которых 33 найдены у человека. CD9 играет роль в клеточной миграции и адгезии.

CD151 — мембранный белок, гликопротеин из надсемейства тетраспанинов, продукт гена человека TSPAN24. Белок определяет группу крови системы RAPH.

CD81 — мембранный белок из надсемейства тетраспанинов, продукт гена человека TSPAN28.
Путь Э́нтнера — Ду́дорова, или КДФГ-путь — путь окисления глюкозы, приводящий к образованию из одной молекулы глюкозы двух молекул пирувата, одной молекулы АТФ и двух молекул восстановленных пиридиновых нуклеотидов. Хотя ранее считалось, что он имеет место лишь у небольшого числа грамотрицательных бактерий, в настоящее время установлено, что этот путь распространён в природе чрезвычайно широко и используется различными группами грамположительных и грамотрицательных бактерий, а также некоторыми археями и даже эукариотами.
Метилглиокса́левый шунт — метаболический путь, встречающийся у некоторых бактерий и представляющий собой отличный от гликолитических реакций путь окисления дигидроксиацетонфосфата до пирувата.

Фруктозо-2,6-бисфосфат — регуляторная молекула всех эукариот, аллостерически, влияющая на активность ферментов фосфофруктокиназы 1 (ФФК-1) и фруктозо-1,6-бисфосфатазы (ФБФаза-1). Усиливает гликолиз и подавляет глюконеогенез. Представляет собой эфир фосфорной кислоты и фруктозы.
Фосфофруктокиназа — фермент-киназа, отвечающий за фосфорилирование фруктозо-6-фосфата в процессе гликолиза.

Фосфофруктокиназа-2 (ФФК-2) или фруктозо-2,6-бисфосфатаза (ФБФ-2) — бифункциональный фермент, регулирующий процессы гликолиза и глюконеогенеза в теле человека. Этот белок является димером, в котором каждый мономер, в зависимости от внешних условий, может действовать как киназа или фосфатаза. Регулируется глюкагоном и инсулином.

Гликосо́ма — органелла, окружённая мембраной и содержащая ферменты гликолиза. Термин был введён Скотом и Стиллом в 1968 году, когда они показали, что гликоген, содержащийся в клетке, есть не статичная, а динамичная молекула. Гликосома имеется у нескольких видов протистов, а именно у ряда представителей класса кинетопластид (Kinetoplastea), среди которых есть возбудители таких болезней человека, как сонная болезнь, болезнь Шагаса и лейшманиоз. Органелла окружена одной мембраной и содержит плотный белковый матрикс. Считается, что она произошла от пероксисомы. Эта точка зрения была подтверждена работами по генетике лейшманий (Leishmania). В настоящее время разрабатываются лекарства, действующие на функционирование гликосом.

Коне́чные проду́кты глики́рования — это белки или липиды, которые подверглись гликированию углеводами. Они могут быть одним из факторов старения и развития или осложнения многих дегенеративных заболеваний, таких как диабет, атеросклероз, хроническая болезнь почек и болезнь Альцгеймера.

DC-SIGN — мембранный белок, рецептор, продукт гена CD209. Является C-лектиновым рецептором и присутствует на поверхности макрофагов и дендритных клеток. Макрофагальный DC-SIGN распознаёт маннозо-содержащие углеводы, которые относятся к классу молекулярных паттернов, связанных с патогенами, и, таким образом, белок относится к рецепторам опознавания паттерна. Связывание белка с патогенами активизирует их фагоцитоз. На миелоидных и пре-плазмоцитоидных дендритных клетках DC-SIGN опосредует их слабые взаимодействия с эндотелиальными клетками и активизирует CD4+ T-лимфоциты, а также распознавание молекулярных паттернов, связанных с патогенами.