Мута́ция — стойкое изменение генома. Термин предложен Хуго де Фризом в 1901 году.

Гисто́ны — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5, H2A, H2B, H3, H4. Гистоны H2A, H2B, H3, H4, называемые кóровыми гистонами, формируют нуклеосому, представляющую собой белковую глобулу, вокруг которой накручена нить ДНК. Гистон H1/H5, называемый линкерным гистоном, связывается с внешней стороной нуклеосомы, фиксируя на ней нить ДНК. В хроматине гистоны составляют 25—40 % сухого веса. Благодаря высокому содержанию лизина и аргинина гистоны проявляют сильно оснóвные свойства. Гистоны непосредственно контактируют с ДНК и способны нейтрализовать отрицательный заряд фосфатных групп ДНК за счёт положительных зарядов аминокислотных остатков. Последовательность аминокислот в этих белках является консервативной и практически не различается в организмах различных таксонов. Гистоны присутствуют в ядрах эукариотических клеток; у бактерий гистонов нет, но они выявлены у архей группы Euryarchaea.

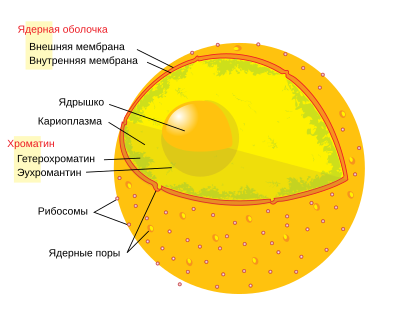
Кле́точное ядро́ — окружённая двумя мембранами важная структура эукариотической клетки. В клетках прокариот ядра нет. В клетках эукариот обычно одно ядро, однако некоторые типы клеток, например, эритроциты млекопитающих, не имеют ядра, а другие содержат несколько ядер.

Плазми́ды — небольшие молекулы ДНК, физически обособленные от хромосом и способные к автономной репликации. Главным образом плазмиды встречаются у бактерий, а также у некоторых архей и эукариот. Чаще всего плазмиды представляют собой двухцепочечные кольцевые молекулы. Несмотря на способность к размножению, плазмиды, как и вирусы, не рассматриваются в качестве живых организмов.
Я́дерный ма́трикс, или я́дерный скеле́т — скелетная структура клеточного ядра, поддерживающая форму и некоторые особенности морфологии ядра. В состав ядерного матрикса входят ядерная ламина, остаточное ядрышко и так называемый диффузный матрикс — сеть филаментов и гранул, соединяющих ядерную ламину с остаточным ядрышком.

Мейо́з, или редукционное деление — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа. В результате мейоза образуются гаметы, споры и другие зародышевые клетки.

Реплика́ция ДНК — процесс создания двух дочерних молекул ДНК на основе родительской молекулы ДНК. Репликацию ДНК осуществляет сложный комплекс, состоящий из 15—20 различных белков-ферментов, называемый реплисомой. С помощью специальных ферментов двойная спираль материнской ДНК расплетается на две нити, на каждой образовавшейся нити достраивается вторая нить, образуя две идентичных дочерних молекулы ДНК, которые затем скручиваются в отдельные спирали. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение.

Цитозин — органическое соединение, азотистое основание, производное пиримидина. С рибозой образует нуклеозид цитидин, входит в состав нуклеотидов ДНК и РНК. Во время репликации и транскрипции по принципу комплементарности образует три водородных связи с гуанином.

ДНК-метилтрансфера́зы (ДНК-метилазы, англ. DNA methyltransferase, DNA MTase, DNMT) — группа ферментов, катализирующих метилирование нуклеотидных остатков в составе ДНК. Активность метилтрансфераз, заключающаяся в переносе метильных (CH3—) групп на азотистое основание цитозин в составе ДНК, ведет к изменению свойств ДНК, при этом изменяется активность, функции соответствующих генов, а также пространственная структура нуклеиновой кислоты (конформация).
Нуклеопротеиды — комплексы нуклеиновых кислот с белками.

ДНК-полимераза — фермент, участвующий в репликации ДНК. Ферменты этого класса катализируют полимеризацию дезоксирибонуклеотидов вдоль цепочки нуклеотидов ДНК, которую фермент «читает» и использует в качестве шаблона. Тип нового нуклеотида определяется по принципу комплементарности с шаблоном, с которого ведётся считывание. Собираемая молекула комплементарна шаблонной моноспирали и идентична второму компоненту двойной спирали.

Белки́ скользя́щего зажима, или скользя́щий зажи́м — белки, которые выполняют функцию усилителя процессивности при репликации ДНК.

Экспериме́нт Мезельсо́на и Ста́ля (англ. Meselson–Stahl experiment — эксперимент, проведённый двумя молекулярными биологами — Мэтью Мезельсоном и Франклином Сталем в 1958 году. Он показал, что репликация ДНК имеет полуконсервативный характер. Это означает, что каждая дочерняя двойная спираль ДНК состоит из одной старой цепи и из одной вновь синтезированной цепи.
Археогенетика — область исследований молекулярной генетики, в которой методы популяционной генетики применяются к изучению истории человечества. Автором термина «археогенетика» является британский археолог Колин Ренфрю.
Цитомегаловирус (человека), или герпесвирус человека тип 5 — вид герпесвирусов, принадлежащих к роду цитомегаловирус (Cytomegalovirus), способных инфицировать человека наряду с другими видами герпеса человека, такими как HSV-1, HSV-2 и вирус Эпштейна — Барр. Вирус был открыт в 1956 году, в 1979 году зарегистрирован в Международном комитете по таксономии вирусов (ICTV), в 2016 году сменил научное название, как и все герпесвирусы.

Пласто́м — генетический материал, содержащийся в пластидах растительной клетки.

Нуклеоли́н — белок, имеющийся у высших эукариот. У человека он кодируется геном NCL, расположенным на 2-й хромосоме в локусе 2q37.1.
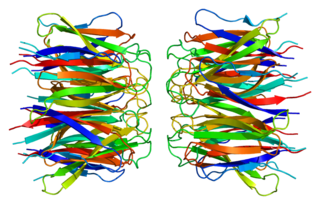
Нуклеофозми́н — ядрышковый белок, у человека кодируется геном NPM1, локализованным на 5-й хромосоме. Нуклеофозмин перемещается между ядром и цитоплазмой и действует как многофункциональный шаперон нуклеиновых кислот, принимающий участие в таких процессах, как биогенез рибосом, ремоделирование хроматина, регуляция митоза, поддержание стабильности генома, репарация ДНК и транскрипция. Нарушения в работе нуклеофозмина могут приводить к развитию злокачественных новообразований и других заболеваний; в частности, мутации, затрагивающие его ген, приводят к развитию острого миелоидного лейкоза.
Я́дерные тельца́ — субкомпартменты внутри ядра, не окружённые мембранами, но представляющие собой отдельные, морфологически различимые комплексы белков и РНК. К числу ядерных телец относят ядрышко, тельце Кахаля и другие немембранные структуры. В основе биогенеза ядерных телец лежат одни и те же общие принципы, такие как способность к формированию de novo, самоорганизация, а также роль РНК как структурного элемента. Контроль биогенеза ядерных телец необходим для правильного изменения архитектуры ядра в ходе клеточного цикла и лежит в основе ответа клетки на внутри- и внеклеточные стимулы. Многие ядерные тельца осуществляют специфические функции — например, синтез и процессинг пре-рибосомных РНК в ядрышке, накопление и сборку компонентов сплайсосом в ядерных спеклах или накопление молекул РНК в параспеклах. Механизмы, которые обеспечивают выполнение ядерными тельцами этих функций, очень разнообразны. В некоторых случаях ядерное тельце может служить местом протекания определённых процессов, например, транскрипции. В других случаях ядерные тельца, по-видимому, опосредованно регулируют локальные концентрации своих компонентов в нуклеоплазме. Хотя большинство ядерных телец имеет сферическую форму, большинство из них можно идентифицировать по уникальной морфологии, которая выявляется при помощи электронной микроскопии, и по расположению в ядре. Подобно цитоплазматическим органеллам, ядерные тельца содержат специфический набор белков, которые определяют их структуру на молекулярном уровне.

Я́дерные спе́клы, или спе́клы, или B-снурпосо́мы, или компа́ртменты фа́кторов спла́йсинга, или доме́ны SC-35, или кла́стеры интерхромати́новых гра́нул — ядерные тельца, регулирующие сплайсинг. В ядерных спеклах находятся факторы сплайсинга и малые ядерные рибонуклеопротеины. Изменения в белковом составе и функционировании ядерных спекл приводят к изменениям в альтернативном сплайсинге пре-мРНК, поэтому считается, что ключевой функцией ядерных спекл является регуляция доступности факторов сплайсинга в сайтах транскрипции. Недавние исследования показали, что в ядерных спеклах также находятся белки, задействованные в регуляции локализации хромосом, модификации хроматина, транскрипции, процессинге 3'-концов транскриптов, модификации мРНК, белки, полностью покрывающие мРНК, и мРНК-рибонуклеопротеины, поэтому ядерные спеклы рассматриваются как крупнейшие центры регуляции всех этапов экспрессии ядерных генов.