Агонист — химическое соединение (лиганд), которое при взаимодействии с рецептором изменяет его состояние, приводя к биологическому отклику. Обычные агонисты увеличивают отклик рецептора, обратные агонисты уменьшают его, а антагонисты блокируют действие рецептора.

Антагонист в биохимии и фармакологии — подтип лигандов к клеточным рецепторам. Лиганд, обладающий свойствами антагониста рецепторов — это такой лиганд, который блокирует, снижает или предотвращает вызываемые связыванием агониста с рецептором физиологические эффекты. Сам же он при этом не обязан производить какие-либо физиологические эффекты вследствие своего связывания с рецептором. Таким образом, антагонисты рецепторов имеют сродство (аффинность) к данному конкретному типу рецепторов, но, исходя из строгого определения, не имеют собственной внутренней агонистической активности по отношению к этому рецептору, и их связывание лишь нарушает взаимодействие [конкурентных] полных или частичных агонистов с рецептором и предотвращает или ингибирует их функцию и их физиологические эффекты. В равной степени антагонисты рецепторов предотвращают и воздействие на рецептор обратных агонистов. Антагонисты рецепторов опосредуют свои эффекты благодаря связыванию либо с активным сайтом рецептора, тем же самым, с которым связывается и физиологический эндогенный агонист, либо с аллостерическими сайтами, или же они могут взаимодействовать с рецептором в уникальных участках связывания, которые не являются нормальными участками связывания эндогенных веществ для данного рецептора и не принимают в норме участия в физиологической регуляции активности данного рецептора.

Габоксадол (THIP) — частичный агонист рецепторов ГАМКA и антагонист рецепторов ГАМКС, который предалагался фармацевтической компанией Merck в качестве экспериментального снотворного препарата. Клинические исследования габоксадола были остановлены досрочно в марте 2007 года.

В биохимии и фармакологии лиганд — это химическое соединение, которое образует комплекс с той или иной биомолекулой и производит, вследствие такого связывания, те или иные биохимические, физиологические или фармакологические эффекты. В случае связывания лиганда с белком лиганд обычно является малой сигнальной молекулой, связывающейся со специфическим участком связывания на белке-мишени. В случае связывания лиганда с ДНК лиганд обычно также является малой молекулой или ионом, или белком который связывается с двойной спиралью ДНК.

ГАМКА-рецептор — лиганд-зависимый ионный канал в химических синапсах нервной системы, который тормозит передачу нервного возбуждения и управляется с помощью ГАМК. Это, наряду с ГАМКС-рецептором, один из двух ионотропных ГАМК-рецепторов, отвечающий за реакцию организма на гамма-аминомасляную кислоту. Помимо места, связывающего ГАМК, рецепторный комплекс содержит аллостерические сегменты, способные связывать бензодиазепины, барбитураты, этанол, фуросемид, нейростероиды и пикротоксин.
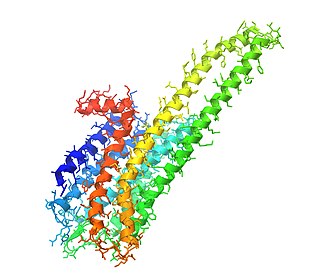
5-HT2A-рецептор у млекопитающих является одним из подтипов 5-HT2-рецепторов, подсемейства серотониновых рецепторов. Он является метаботропным G-белок-сопряжённым рецептором. Рецептор этого подтипа (5-HT2A) является основным возбуждающим подтипом рецепторов среди всех G-белок-сопряжённых подтипов рецепторов для серотонина (5-HT). Однако рецепторы подтипа 5-HT2A способны, наоборот, оказывать ингибирующее, тормозящее воздействие в некоторых областях мозга, таких, как зрительная кора и орбитофронтальная кора.

Опио́идные реце́пторы (опиатные рецепторы) — разновидность рецепторов нервной системы, относящихся к рецепторам, сопряжённым с G белком. Основная их функция в организме — регулирование болевых ощущений. В настоящее время различают четыре основные группы опиоидных рецепторов: μ- (мю), δ- (дельта), κ- (каппа) и ноцицептиновые (ORL1) рецепторы. Они связываются как с эндогенными (вырабатываемые в организме), так и с экзогенными (поступающими извне) опиоидными лигандами. Опиатные рецепторы широко распространены в головном, спинном мозге, а также в желудочно-кишечном тракте и других органах.

Никотиновый ацетилхолиновый рецептор (н-холинорецептор, nACh-receptor) — подвид ацетилхолиновых рецепторов, который обеспечивает передачу нервного импульса через синапсы и активируется ацетилхолином, а также никотином. Этот рецептор вместе с ГАМКA-, ГАМКC- глициновым и 5-HT3-рецепторами образует семейство лиганд-зависимых ионных каналов с цистеиновой петлёй.
5-HT1A-рецептор — подтип серотониновых рецепторов подсемейства 5-HT1-рецепторов. Эндогенным лигандом-агонистом для них является моноаминовый нейромедиатор серотонин (5-гидрокситриптамин, 5-HT). Рецепторы этого подтипа относятся к семейству трансмембранных метаботропных G-белок-связанных рецепторов и связаны с гетеротримерным ингибиторным G-белком, так называемым Gi / Go. Этот подтип рецепторов опосредует тормозящую нейротрансмиссию. Ген, кодирующий белок этого рецептора у человека, обозначается HTR1A.

5-HT₇-рецептор — это один из подтипов серотониновых рецепторов, относящийся к семейству метаботропных G-белок-связанных рецепторов на мембране клетки. Он активируется нейромедиатором серотонином. Рецепторы подтипа 5-HT₇ связаны с гетеротримерным G-белком Gs, который стимулирует образование внутриклеточных сигнальных молекул — молекул цАМФ. Он экспрессируется в различных тканях человека, в особенности в мозге, в желудочно-кишечном тракте и в кровеносных сосудах. В последнее время 5-HT₇-рецептор является мишенью для разработки новых лекарств, предназначенных для лечения различных заболеваний, в частности, лекарств для лечения депрессий, то есть антидепрессантов. Рецепторы подтипа 5-HT₇ кодируются геном HTR7, который у человека транскрипционируется путём альтернативного сплайсинга в 3 различных сплайс-варианта — 5-HT₇(a), 5-HT₇(b), 5-HT₇(d). Вариант 5-HT₇(c) у человека отсутствует, зато имеется у крыс, у которых отсутствует 5-HT₇(d).

Внутренняя агонистическая активность или «внутренняя активность», «агонистическая активность», или «рецепторная эффективность», «эффективность по отношению к рецептору», коротко называемая «эффективность» в контексте фармакологии — термин, которым обозначают меру относительной способности комплекса лиганда с рецептором производить максимальный физиологический ответ.

В фармакологии термин части́чные агони́сты применяется по отношению к лекарствам и химическим соединениям, которые являются лигандами для конкретного подтипа клеточных рецепторов и способны активировать рецептор, то есть переводить его в активную пространственную конфигурацию, но с меньшей вероятностью, чем эндогенный агонист тех же рецепторов, рецепторная эффективность которого принимается за 100 % и который рассматривается, таким образом, как истинный полный агонист. Другими словами, внутренняя агонистическая активность частичного агониста по определению всегда больше 0 %, но меньше 100 %.
Конкурентные антагонисты — подтип лигандов-антагонистов рецепторов, которые обратимо связываются с рецепторами в том же самом участке связывания, что и физиологический эндогенный лиганд-агонист этого рецептора, но не вызывают активации рецептора. Физиологические агонисты и конкурентные антагонисты в этом случае «конкурируют» за связывание с одним и тем же участком связывания рецепторов. После того, как конкурентный антагонист свяжется с участком связывания рецептора, он предотвращает связывание с ним же агониста. Однако конкурентный антагонист не может ни «вытеснить» уже связавшийся с рецептором агонист из связи, ни предотвратить воздействие уже связавшегося агониста на клетку. Конечный результат конкуренции агонистов и антагонистов — и таким образом конечный уровень активности рецепторной системы — определяется соотношением молярных концентраций, относительным сродством к рецепторам и соотношением относительной внутренней агонистической активности агонистов и антагонистов. Поскольку высокие концентрации конкурентного антагониста повышают процент занятости рецепторов этим антагонистом, для достижения того же самого процента занятости рецепторов агонистом в этих условиях — и получения того же самого физиологического ответа — потребуются более высокие концентрации агониста, и наоборот — при более высоких концентрациях агониста требуется больше конкурентного антагониста для функциональной «блокады» рецепторов. В функциональных исследованиях конкурентные антагонисты вызывают параллельный сдвиг кривой «доза агониста-эффект» вправо, без изменения максимальной величины физиологического ответа.
Термин «неконкурентный антагонизм» используется для описания двух разных феноменов: в одном случае неконкурентный антагонист связывается с ортостерическим участком связывания лигандов на рецепторе, а в другом случае он связывается с аллостерическим участком связывания лигандов на рецепторе. И хотя механизм антагонистического действия различен в обоих случаях, они оба называются «неконкурентным антагонизмом», поскольку конечный результат воздействия антагониста в обоих случаях функционально очень похож. В отличие от конкурентных антагонистов, которые конкурируют с агонистами за занятость рецепторов и сдвигают кривую доза-эффект вправо, влияя на количество агониста, необходимое для получения максимального физиологического ответа, но никак не влияют на саму величину максимального физиологического ответа, неконкурентные антагонисты уменьшают величину максимального физиологического ответа, который может быть получен при любом сколь угодно большом количестве агониста. Это свойство и даёт им название «неконкурентные антагонисты», поскольку их эффект не может быть «уничтожен», обнулен или скомпенсирован увеличением количества агониста, сколь бы велико это увеличение ни было. В биологических системах, предназначенных для изучения влияния тех или иных антагонистов на рецепторы, неконкурентные антагонисты вызывают уменьшение «плато», и, в некоторых случаях, также сдвиг кривой вправо. Сдвиг кривой вправо происходит вследствие наличия во многих биологических рецепторных системах так называемого «рецепторного резерва», и ингибирование агонистического ответа под влиянием неконкурентного антагониста происходит только тогда, когда истощится (израсходуется) этот рецепторный резерв.
«Молчаливые антагонисты», или нейтральные антагонисты — это такие конкурентные антагонисты данного типа рецепторов, которые имеют строго нулевую внутреннюю агонистическую активность, то есть нулевую способность активировать рецептор, но также и не препятствуют конституциональной внутренней активности рецептора, не снижают её и не обладают собственными, отличными от блокирования связывания рецептора с агонистом, физиологическими эффектами по отношению к данной рецепторной системе. В некотором смысле именно «молчаливые антагонисты» являются «истинными», «настоящими» антагонистами, в первоначальном значении слова.
Необратимые антагонисты ковалентно связываются с рецептором-мишенью, необратимо меняя его пространственную конфигурацию и тем самым необратимо инактивируют его. Необратимые антагонисты, как правило, не могут быть ферментативно удалены из связи с рецептором. Таким образом, продолжительность физиологического воздействия необратимого антагониста определяется не традиционной кинетикой связывания рецептора с лигандом, а скоростью оборота рецепторов — скоростью процесса физиологического «выведения из строя» и удаления с поверхности мембраны клетки старых, «деградировавших» от времени рецепторов и скоростью биосинтеза клеткой и выведения на поверхность клеточной мембраны новых рецепторов взамен старых, деградировавших. Примером необратимого антагониста α-адренорецепторов является феноксибензамин, который ковалентно и необратимо связывается с α-адренорецепторами, предотвращая тем самым связывание с ними адреналина и норадреналина. Инактивация рецепторов необратимым агонистом обычно приводит к снижению или уменьшению максимально возможного физиологического ответа на максимальную агонистическую стимуляцию. Помимо этого, в системах, в которых есть рецепторный резерв, также может наблюдаться сдвиг кривой «доза-эффект» вправо, подобный сдвигу кривой вправо, наблюдаемому при воздействии конкурентных антагонистов. Отмывание культуры клеток, которые подверглись воздействию антагониста, от остатков антагониста, обычно позволяет различить воздействие неконкурентного антагониста от воздействия необратимого антагониста, поскольку действие неконкурентных антагонистов кратковременно и обратимо, и после отмывания клеток от антагониста эффективность воздействия на них агониста восстанавливается, чего не происходит при воздействии необратимых антагонистов.
В фармакологии, физиологии и биохимии, термин эндогенный агонист по отношению к определённому подтипу клеточных рецепторов обозначает химическое соединение, которое отвечает следующим трём условиям:
- производится в самом организме или ткани в нормальных физиологических условиях, то есть является эндогенным веществом — например, нейромедиатором, цитокином или гормоном;
- является лигандом для данного подтипа рецепторов, то есть обладает способностью связываться с ортостерическим сайтом связывания данного подтипа рецепторов;
- обладает способностью после связывания активировать этот рецептор, изменяя его пространственную конфигурацию, что приводит к вызыванию соответствующего биохимического или физиологического ответа.

H3-гистами́новый реце́птор (сокр. H3), также H3-реце́птор — интегральный мембранный белок, один из 4-х видов гистаминовых рецепторов, принадлежит к суперсемейству родопсинподобных рецепторов, связанных с G-белком. Активируется посредством связывания гистамина. У человека ген, кодирующий данный белок HRH3, локализован на длинном плече (q-плече) 20-й хромосомы. Белок состоит из последовательности 445 аминокислот и имеет молекулярную массу равную 48671 Да.

Лиганд-зависимые ионные каналы, лиганд-управляемые или лиганд-активируемые ионные каналы — также обычно называемые ионотропными рецепторами, представляют собой группу трансмембранных белков ионных каналов, которые позволяют ионам, например, Na+, K+, Ca2+ и/или Cl-, проходить через биологическую мембрану, посредством изменения конформации (открытия) в ответ на связывание химического мессенджера (то есть лиганда), такого как, например, молекулы нейромедиатора.

В фармакологии и биохимии аллостерические модуляторы представляют собой группу веществ, которые связываются с рецептором, чтобы изменить реакцию этого рецептора на стимул. Некоторые из них, такие как бензодиазепины, являются наркотиками. Сайт, с которым связывается аллостерический модулятор, отличается от того, с которым связывался бы эндогенный агонист рецептора. Модуляторы и агонисты могут быть названы лигандами рецепторов.

