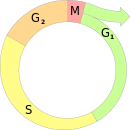
Хромосо́мы — нуклеопротеидные структуры в ядре эукариотической клетки, в которых сосредоточена бо́льшая часть наследственной информации и которые предназначены для её хранения, реализации и передачи. Хромосомы чётко различимы в световом микроскопе только в период митотического или мейотического деления клетки. Набор всех хромосом клетки, называемый кариотипом, является видоспецифичным признаком, для которого характерен относительно низкий уровень индивидуальной изменчивости.

Мито́з, или кариокине́з — непрямое деление клетки, наиболее распространённый способ размножения эукариотических клеток. Биологическое значение митоза состоит в строго одинаковом распределении хромосом между дочерними ядрами, что обеспечивает образование генетически одинаковых дочерних клеток и сохраняет преемственность в ряду клеточных поколений. Перед делением число хромосом в клетке увеличивается в два раза продольным разделением на две части каждой из них, поэтому в каждую из дочерних клеток переходит столько же хромосом, сколько их было в родительской клетке.

Мейо́з, или редукционное деление — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа. В результате мейоза образуются гаметы, споры и другие зародышевые клетки.

Веретено́ деле́ния — динамичная структура, которая образуется в митозе и мейозе для обеспечения сегрегации (отделения) хромосом и деления клетки. Типичное веретено является биполярным — между двумя полюсами образуется веретенообразная система микротрубочек. Микротрубочки веретена присоединяются к кинетохорам хроматид в области центромер и обеспечивают движение хромосом по направлению к полюсам.

Центромера — участок хромосомы, который связывает сестринские хроматиды, играет важную роль в процессе деления клеточного ядра и участвует в контроле экспрессии генов. Характеризуется специфическими последовательностью нуклеотидов и структурой.

Клеточный центр, или центросома — немембранная органелла в клетках эукариот, состоит из двух центриолей и перицентриолярного материала. Является главным центром организации микротрубочек (ЦОМТ) эукариотической клетки, играет важнейшую роль в клеточном делении, участвуя в формировании веретена деления. Из центросомы образуются реснички и жгутики. Центросомы характерны для клеток животных, их нет у высших растений, у низших грибов и некоторых простейших. Ряд наследственных заболеваний человека вызван мутациями в генах, кодирующих центросомные белки.

Метафа́за — фаза митотического деления эукариотических клеток, начало которой знаменует выход хромосом в экваториальную плоскость клетки.

Профа́за — самая первая фаза митоза, признаком которой является появление в ядре конденсированных хромосом.

Анафа́за — фаза митотического деления эукариотических клеток, в которой сестринские хроматиды синхронно расходятся с образованием двух дочерних хромосом, которые неспешно растаскиваются к противоположным полюсам веретена деления. В ходе анафазы кинетохорные микротрубочки укорачиваются, а полюса удаляются друг от друга, таким образом, оба процесса вносят свой вклад в расхождение хроматид.

Телофа́за — фаза митотического деления эукариотических клеток, во время которой два набора дочерних хромосом достигают полюсов веретена деления и деконденсируются. Начинается сборка ядерной оболочки вокруг каждого набора хромосом. Разделение цитоплазмы достигается путём сокращения сократительного кольца (цитокинез).

Кинетохор — белковая структура на хромосоме, к которой крепятся волокна веретена деления во время деления клетки. Кинетохоры играют важнейшую роль при сегрегации хромосом для последующего разделения родительской клетки на две дочерние.
Ана-телофазный анализ — генетический тест, основанный на визуальном учёте хромосомных аберраций на стадии анафазы и телофазы митотического цикла клетки. Ана- телофазный анализ — простой, экономичный метод, который не требует знания кариотипа и идентификации хромосом. Он позволяет выявить лишь определенные типы хромосомных аберраций, но его чувствительность вполне достаточна для заключения «мутагенен» или «не мутагенен» фактор. Ана- телофазный анализ является достаточно чувствительным, корректным и удобным на первом этапе экотоксикогенетического исследования.

Препрофа́за — дополнительная фаза митоза, имеющаяся у растительных клеток и отсутствующая у других эукариот — животных и грибов. Она предшествует профазе и включает два основных события:
- Образование препрофазной ленты, густого кольца из микротрубочек, расположенного под плазматической мембраной;
- Начало скопления микротрубочек около ядерной оболочки.

Препрофа́зная ле́нта — кольцо из микротрубочек, обнаруженное в растительных клетках, подвергающихся делению и вступивших в фазу препрофазы клеточного цикла клеток растений. Не считая фрагмосомы, препрофазная лента — первая видимая под микроскопом структура в растительных клетках, вступивших в митоз.
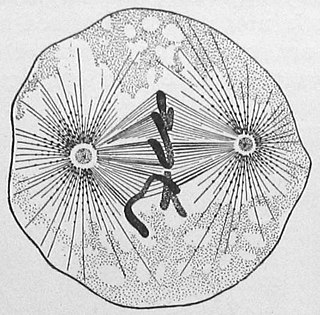
Экваториальная (метафазная) пластинка — расположение хромосом во время одной из стадий деления ядра клетки (метафазы), во время которой они располагаются по экватору ядерного веретена и при рассматривании сбоку при малых увеличениях производят — из-за невозможности рассмотреть отдельные хромосомы в плоскости экватора в это время — впечатление хроматиновой пластинки, лежащей по экватору, то есть на равном расстоянии от обоих полюсов клетки. После образования экваториальной пластинки начинается процесс разъединения сестринских хромосом и расхождения их к противоположным полюсам (анафаза).
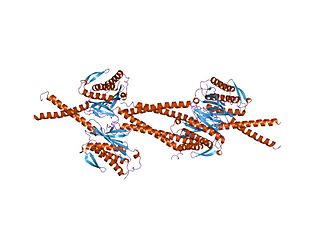
Mad1 — белок дрожжей и других эукариот, который участвует в прохождении контрольной точки сборки веретена деления. Эта контрольная точка контролирует прикрепление микротрубочек к хромосомам и препятствует вхождению клетки в анафазу до тех пор, пока веретено деления не будет полностью собрано. Название Mad дано за то, что у клеток, мутантных по этому белку, наблюдается дефектность блокировки митоза при деполимеризации микротрубочек. MAD1 рекрутирует ингибитор анафазы Mad2 к свободным кинетохорам, не соединившимся с микротрубочками веретена, и имеет важное значение для образования комплекса Mad2-Cdc20 in vivo, но не in vitro. In vivo Mad1 действует как конкурентный ингибитор комплекса Mad2-Cdc20. Mad1 фосфорилируется киназой Mps1, что вместе с другими процессами приводит к формированию комплекса митотической контрольной точки. Таким образом он ингибирует активность комплекса стимуляции анафазы/циклосомы (APC/C). Гомологи Mad1 эволюционно консервативны у эукариот от дрожжей до млекопитающих. Регулируя клеточный цикл и расхождение хромосом, а также выполняя некоторые интерфазные функции, Mad1 оказывается вовлечённым в развитие многих опухолей и генетических заболеваний.

Киназа Aurora B — белок, прикрепляющий митотическое веретено деления к центромере.

Mad2 является важным белком контрольной точки веретена деления. Система контрольной точки веретена деления является системой регулирования, сдерживающей прогрессирование перехода от метафазы к анафазе. Ген Mad2 был впервые выявлен в дрожжах S. cerevisiae в скрининге для генов, мутированных для придания им чувствительности к ядам микротрубочек. Человеческие ортологи Mad2 были впервые выделены в поисках человеческих кДНК, которые спасают микротрубочки ядочувствительного штамма дрожжей, в котором отсутствовал белок, связывающий кинетохор. Было показано, что белок был представлен в одиноких кинетохорах, и исследование ингибирования антител показало, что он имеет важное значение для выполнения блока перехода от метафазы к анафазе в ответ на отравление микротрубочек нокодазолом. После выделения ортолога Xenopus Laevis , подобного человеческим последовательностям, появилась возможность охарактеризовать митотическую контрольную точку в экстрактах яиц.

Полярное тельце веретена (ПТВ) — центр организации микротрубочек, грибной эквивалент центросомы клеток животных. В отличие от центросомы в ПТВ нет центриолей. У дрожжей S. cerevisiae под электронным микроскопом оно выглядит как электронно-плотная многослойная структура, встроенная в оболочку ядра. Помимо основной функции, полярное тельце веретена опосредованно участвует в сегрегации хромосом, расположении ядер в клетке, кариогамии и ориентации веретена деления. Кроме того, оно является местом восприятия сигналов MEN пути, и, возможно участвует в формировании стенки сумки споры S. cerevisiae.

Контрольная точка веретена, также известная как переход от метафазы к анафазе, контрольная точка сборки веретена (SAC), контрольная точка метафазы или митотическая контрольная точка, представляет собой контрольную точку клеточного цикла во время митоза или мейоза, которая предотвращает разделение дуплицированных хромосом (анафазу) до тех пор, пока каждая хромосома не будет должным образом прикреплена к веретену. Для достижения правильной сегрегации две кинетохоры на сестринских хроматидах должны быть прикреплены к противоположным полюсам веретена. Только такой способ прикрепления гарантирует, что каждая дочерняя клетка получит одну копию хромосомы. Определяющей биохимической особенностью этой контрольной точки является стимуляция комплекса, способствующего анафазе, комплексами M-фазы циклин-CDK, что, в свою очередь, вызывает протеолитическую деструкцию циклинов и белков, удерживающих вместе сестринские хроматиды.