
Гисто́ны — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5, H2A, H2B, H3, H4. Гистоны H2A, H2B, H3, H4, называемые кóровыми гистонами, формируют нуклеосому, представляющую собой белковую глобулу, вокруг которой накручена нить ДНК. Гистон H1/H5, называемый линкерным гистоном, связывается с внешней стороной нуклеосомы, фиксируя на ней нить ДНК. В хроматине гистоны составляют 25—40 % сухого веса. Благодаря высокому содержанию лизина и аргинина гистоны проявляют сильно оснóвные свойства. Гистоны непосредственно контактируют с ДНК и способны нейтрализовать отрицательный заряд фосфатных групп ДНК за счёт положительных зарядов аминокислотных остатков. Последовательность аминокислот в этих белках является консервативной и практически не различается в организмах различных таксонов. Гистоны присутствуют в ядрах эукариотических клеток; у бактерий гистонов нет, но они выявлены у архей группы Euryarchaea.
Нуклеосома — это структурная часть хромосомы, образованная совместной упаковкой нити ДНК с гистоновыми белками H2А, H2B, H3 и H4. Последовательность нуклеосом, соединенная гистоновым белком H1, формирует нуклеофиламент, или иначе нуклеосомную нить.
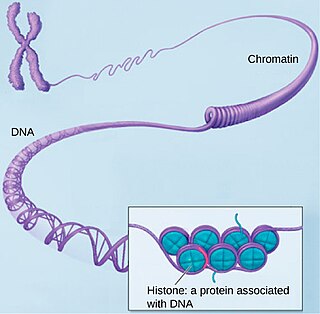
Хроматин — нуклеопротеид, составляющий основу хромосом. Состоит из ДНК и белков. Хроматин находится внутри ядра клеток эукариот и архей, имеющих гистоны. В более широком смысле хроматином иногда называют также и вещество нуклеоида у бактерий.

Некодирующие РНК — молекулы РНК, которые не транслируются в белки. Ранее использовавшийся синоним — «малые РНК» — вышел из употребления, так как некоторые некодирующие РНК могут быть очень большими, например, Xist.
Белки группы polycomb — это семейство белков, которые способны ремоделировать хроматин. Эти белки-регуляторы были впервые описаны у дрозофил, где они подавляют гомеозисные гены, контролирующие индивидуальные отличия сегментов развивающегося эмбриона.

Эухромати́н, также активный или «открытый хромати́н» — участки хроматина, которые представляет собой неплотную и легко упакованную форму, обогащённую генами и часто находящуюся в состоянии активной транскрипции. Эухроматин отличается от гетерохроматина, который плотно упакован и менее доступен для транскрипции. Около 92 % генома человека является эухроматичным.
Белки группы Trithorax — это белки, регулирующие структуру хроматина и участвующие в поддержании экспрессии генов. Это семейство белков гетерогенно, входящие в него белки образуют сложные белковые комплексы, способные связываться с хроматином. Так, функция самого белка Trithorax в составе комплекса TAC1 — поддержание локального открытого состояния хроматина путём монометилирования лизина-4 гистона H3 (H3K4me1), что сохраняет активность окрестных генов в ряду клеточных поколений. Действие этой гистоновой метки усиливается ацетилированием лизина-27 гистона H3 (H3K27ac), привносимой тем же комплексом TAC1. В дополнение к этому некоторые белки группы Trithorax ремоделируют хроматин, используя энергию АТФ для активации нуклеосом.
Энхансер — небольшой участок ДНК, который после связывания с ним факторов транскрипции стимулирует транскрипцию с основных промоторов гена или группы генов. Энхансеры не обязательно находятся в непосредственной близости от генов, активность которых они регулируют, и даже не обязательно располагаются с ними на одной хромосоме. Энхансеры могут располагаться как в 5'-, так и в 3'-положении относительно матричной цепи регулируемого гена и в любой ориентации к ней. Энхансеры также могут находиться внутри интронов. Тем не менее для работы энхансера необходим его физический контакт с промотором, который осуществляется за счёт «выпетливания» ДНК между энхансером и промотором. Молекулярный механизм действия энхансера заключается в том, что он благодаря собранному на нём белковому комплексу привлекает РНК-полимеразу II и кофакторы транскрипции в область промотора.
Эпигенетика — раздел генетики. Эпигенетика изучает наследуемые изменения активности генов во время роста и деления клеток — изменения синтеза белков, вызванных механизмами, не изменяющими последовательность нуклеотидов в ДНК. Эпигенетические изменения сохраняются в ряде митотических делений соматических клеток, а также могут передаваться следующим поколениям. Регуляторы синтеза белка — метилирование и деметилирование ДНК, ацетилирование и деацетилирование гистонов, фосфорилирование и дефосфорилирование транскрипционных факторов и другие внутриклеточные механизмы.

Когези́н — это мультибелковый комплекс, который регулирует процесс разделения сестринских хроматид в ходе деления клетки.

Откры́тый хромати́н — небольшие участки хроматина, свободные от нуклеосом. Посадке нуклеосом, как правило, препятствуют связанные с хроматином белковые факторы, узнающие определённые последовательности ДНК. К числу таких белков относятся транскрипционные факторы, ДНК- или РНК-полимеразы. Открытый хроматин часто совпадает с цис-регуляторными последовательностями, а именно: промоторами, энхансерами, инсуляторами, сайленсерами, участками начала репликации ДНК. Размер открытых участков хроматина обычно составляет несколько сотен пар нуклеотидов, в среднем около 300 п.н.
Семейство белков BET — семейство белков, содержащих два тандемных бромодомена на N-конце молекулы и экстратерминальный домен. Белки семейства BET являются глобальными корегуляторами транскрипции. Они связывают ε-аминоацетильные группы остатков лизина в гистонах и привлекают в этот комплекс белки ремоделирования хроматина, соответственно они могут выступать в роли коактиваторов или корепрессоров транскрипции в зависимости от молекулярного контекста. Бромодомены белков семейства BET характеризуются относительно низкой аффинностью к одиночным остаткам ацетиллизина к полипептидах и гораздо большей к участкам со множественными сайтами ацетилирования.

CHD3 — фермент, кодируемый у человека геном CHD3.

ATB1 — белок, кодируемый у человека геном SATB1.

Ассоциированный с метастазами белок MTA2 — белок, кодируемый у человека геном MTA2 .

Cas9 — это управляемая при помощи РНК-гидов эндонуклеаза, связанная с адаптивной иммунной системой CRISPR у ряда бактерий, в частности Streptococcus pyogenes. S. pyogenes использует Cas9 для запоминания, последующей проверки и разрезания чужеродной ДНК, например, ДНК бактериофагов или плазмид.
CAF-1 представляет собой комплекс, состоящий из трёх субъединиц: Chaf1a (p150), Chaf1b (p60) и p50. Этот комплекс переносит тетрамер (четвёрку) гистонов H3/H4 на реплицирующуюся ДНК. CAF-1 участвует в создании клеточной эпигенетической памяти, необходимой для поддержания индивидуальных черт (дифференцировки) соматических клеток.
Регуляция транскрипции — совокупность процессов в клетке, посредством которых осуществляется контроль за транскрипцией — синтезом РНК на матрице ДНК — одним из этапов экспрессии генов. Транскрипционная активность гена может контролироваться более чем одним механизмом. Эти механизмы различаются у прокариот и эукариот.
ATAC-seq — метод для полногеномного оценивания степени открытости хроматина. Метод появился в 2013 году как альтернатива MNase-seq, FAIRE-Seq и DNase-seq. По сравнению с DNase-seq и MNase-seq ATAC-seq является более быстрым и чувствительным методом анализа эпигенома.
Эпигено́мика — раздел молекулярной биологии, изучающий совокупность эпигенетических модификаций генетического материала клетки с помощью высокопроизводительных методов. Эпигеномика аналогична геномике и протеомике, которые изучают геном и протеом клетки, соответственно.