
Рибонуклеи́новая кислота́ (РНК) — одна из трёх основных макромолекул, которые содержатся в клетках всех живых организмов и играют важную роль в кодировании, прочтении, регуляции и экспрессии генов.
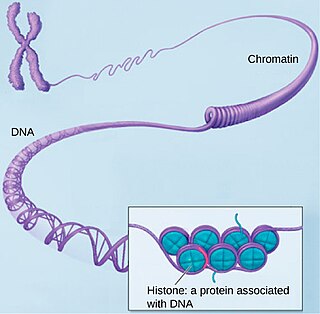
Хроматин — нуклеопротеид, составляющий основу хромосом. Состоит из ДНК и белков. Хроматин находится внутри ядра клеток эукариот и архей, имеющих гистоны. В более широком смысле хроматином иногда называют также и вещество нуклеоида у бактерий.
Гено́м — совокупность наследственного материала, заключённого в клетке организма. Геном содержит биологическую информацию, необходимую для построения и поддержания организма. Большинство геномов, в том числе геном человека и геномы всех остальных клеточных форм жизни, построено из ДНК, однако некоторые вирусы имеют геномы из РНК.

Транспозоны — участки ДНК организмов, способные к передвижению (транспозиции) и размножению в пределах генома. Транспозоны также известны под названием «прыгающие гены» и являются примерами мобильных генетических элементов.
Вирусологическая теория эволюции — эволюционная теория, считающая главным фактором наследственной изменчивости не радиоактивность или другие факторы, а заражение вирусом, изменяющим наследственность заражённого организма.

ДНК-ма́ркеры (ДНК-маркёры), или молекулярно-генетические маркеры — полиморфный признак, выявляемый методами молекулярной биологии на уровне нуклеотидной последовательности ДНК для определённого гена или для любого другого участка хромосомы при сравнении генотипов различных особей, пород, сортов, линий.

Геном человека — совокупность наследственного материала, заключённого в клетке человека. Согласно этому определению человеческий геном состоит из 23 пар хромосом, находящихся в ядре, а также множества копий митохондриальной ДНК. Существует и другое определение генома, в котором под геномом подразумевают совокупность генетического материала гаплоидного набора хромосом. Когда говорят о размере генома человека, то имеют в виду данный вариант определения генома. Так, двадцать две аутосомы, две половые хромосомы Х и Y, а также митохондриальная ДНК человека содержат вместе 3 099 734 149 пар оснований.
Некодирующая ДНК или Мусорная ДНК — части геномной ДНК организмов, которые не кодируют последовательности белков. Некоторые некодирующие ДНК переводятся в функциональные некодирующие РНК-молекулы. Другие функции некодирующей ДНК включают регуляцию последовательностей кодирующих белки, центромер и теломер. Термин «мусорная ДНК» стал популярным в 1960-х. В соответствии с T. Ryan Gregory, геномным биологом, первое явное обсуждение природы «мусорной» ДНК было сделано David Comings в 1972 году и он применил этот термин ко всем некодирующим ДНК. Термин был формализован Сусуму Оно в 1972 году, который заметил, что генетический груз нейтральных мутаций находится на верхнем пределе значений для функционирующих локусов, которые могли быть ожидаемыми исходя из типичной частоты мутаций. Сусуму предсказал, что геномы млекопитающих не могут содержать более чем 30 000 локусов из-за давления естественного отбора, так как «стоимость» мутационной нагрузки вызвала бы неизбежное снижение приспособленности, и в конечном счете вымирание. Этот прогноз остается верным, геном человека содержит приблизительно 20 000 генов. Другим подтверждением теории Оно служит наблюдение, что даже близкородственные виды могут иметь очень разные по размеру геномы, которое окрестили C-парадокс в 1971 году.
piРНК — наиболее крупный класс малых некодирующих РНК, экспрессируемых в клетках животных; они обнаружены в комплексах с белками семейства Piwi, за что и получили своё название. piРНК обычно длиннее микроРНК и малых интерферирующих РНК и имеют длину 26—32 нуклеотида, кроме того, в отличие от микроРНК, они не так консервативны. Белки Piwi относятся к большой группе белков Argonaute и экспрессируются почти исключительно в клетках зародышевой линии; они необходимы для поддержания стволовых клеток зародышевой линии, сперматогенеза и репрессии мобильных элементов. Комплексы Piwi с piРНК не только задействованы в сайленсинге ретротранспозонов и других генетических элементов на пост-трансляционном уровне, но имеют и некоторые другие, в значительной мере ещё неописанные эффекты, например, эпигенетические.
Энхансер — небольшой участок ДНК, который после связывания с ним факторов транскрипции стимулирует транскрипцию с основных промоторов гена или группы генов. Энхансеры не обязательно находятся в непосредственной близости от генов, активность которых они регулируют, и даже не обязательно располагаются с ними на одной хромосоме. Энхансеры могут располагаться как в 5'-, так и в 3'-положении относительно матричной цепи регулируемого гена и в любой ориентации к ней. Энхансеры также могут находиться внутри интронов. Тем не менее для работы энхансера необходим его физический контакт с промотором, который осуществляется за счёт «выпетливания» ДНК между энхансером и промотором. Молекулярный механизм действия энхансера заключается в том, что он благодаря собранному на нём белковому комплексу привлекает РНК-полимеразу II и кофакторы транскрипции в область промотора.
CRISPR — особые локусы бактерий и архей, состоящие из прямых повторяющихся последовательностей, которые разделены уникальными последовательностями (спейсерами). Спейсеры заимствуются из чужеродных генетических элементов, с которыми сталкивалась клетка. РНК, транскрибирующиеся с локусов CRISPR, совместно с ассоциированными белками Cas обеспечивают адаптивный иммунитет за счёт комплементарного связывания РНК с нуклеиновыми кислотами чужеродных элементов и последующего разрушения их белками Cas. Впрочем, к настоящему моменту имеется немало свидетельств участия CRISPR в процессах, не связанных с иммунитетом.
Alu-повтор, Alu-элемент — короткая последовательность ДНК, которая была открыта при обработке ДНК человека рестриктазой Alu. Alu-повтор принадлежит к классу коротких диспергированных повторов (SINEs). Alu-повторы различных типов в большом количестве представлены в геномах приматов. В человеческом геноме они являются одними из самых распространенных элементов. Alu-повтор произошёл от гена, кодирующего 7SL РНК, которая является компонентом сигнал распознающей частицы. Впервые Alu-повтор появился у предков приматов.

Helicos Biosciences — это компания, которая предоставила революционно новую технологию секвенирования единичных молекул, названную true Single Molecule Sequencing (tSMS).
SINEs — короткие последовательности ДНК в геноме эукариот, появившиеся в результате обратной транскрипции коротких молекул РНК, транскрибируемых РНК-полимеразой III: 5S рРНК, тРНК и различные мяРНК.
LINEs — длинные последовательности ДНК в геноме эукариот, представляющие собой ретротранспозоны, не содержащие длинных концевых повторов.

Cas9 — это управляемая при помощи РНК-гидов эндонуклеаза, связанная с адаптивной иммунной системой CRISPR у ряда бактерий, в частности Streptococcus pyogenes. S. pyogenes использует Cas9 для запоминания, последующей проверки и разрезания чужеродной ДНК, например, ДНК бактериофагов или плазмид.
Полинто́ны — крупные ДНК-транспозоны, содержащие гены, гомологичные вирусным белкам; часто встречаются в эукариотических геномах. Эти наиболее крупные и сложно устроенные ДНК-транспозоны были открыты в середине 2000-х годов. Один полинтон может кодировать до 10 различных белков. Название этих мобильных элементов образовано от двух ключевых белков, которые они кодируют: ДНК-полимераза (POLymerase) и интеграза (INTegrase) ретровирусного типа.
Диспергированные повторы — повторяющиеся последовательности нуклеотидов в геноме. Отличаются от тандемных повторов тем, что расположены не последовательно друг за другом, а на расстоянии. Встречаются в эукариотических и прокариотических геномах. Последовательности и количество повторов зависят от конкретного вида и организма.

Ретротранспозоны LTR представляют собой мобильный элемент класса I, характеризующийся наличием длинных концевых повторов (LTR), непосредственно фланкирующих внутреннюю кодирующую область. Как ретротранспозоны, они мобилизуются посредством обратной транскрипции своей мРНК и интеграции вновь созданной кДНК в другое место. Их механизм ретротранспозиции аналогичен ретровирусам, с той разницей, что большинство LTR-ретротранспозонов не образуют инфекционных частиц, которые покидают клетки и, следовательно, реплицируются только внутри своего генома. Те, которые действительно (иногда) образуют вирусоподобные частицы, классифицируются как Ortervirales.