Антитела́, иммуноглобулины — крупные глобулярные белки плазмы крови, выделяющиеся плазматическими клетками иммунной системы и служащие для нейтрализации клеток патогенов и вирусов, а также белковых ядов и некоторых других чужеродных веществ. Каждое антитело распознаёт уникальный элемент патогена, отсутствующий в самом организме, — антиген, а в пределах данного антигена — определённый его участок, эпитоп. Связываясь с антигенами на поверхности патогенов, антитела могут либо непосредственно нейтрализовать их, либо привлекать другие компоненты иммунной системы, такие как система комплемента и фагоциты, чтобы уничтожить чужеродные клетки или вирусные частицы. Антитела — важнейший компонент гуморального специфического иммунитета.

Систе́ма комплеме́нта — комплекс защитных белков, постоянно присутствующих в крови. Это каскадная система протеолитических ферментов, предназначенная для гуморальной защиты организма от действия чужеродных агентов, она участвует в реализации иммунного ответа организма. Является важным компонентом как врождённого, так и приобретённого иммунитета. Выделяют три основных пути активации системы комплемента: классический, альтернативный и лектиновый. Для запуска классического пути комплемента необходима опсонизация чужеродной клетки антителами, а альтернативный и лектиновый пути могут активироваться в отсутствие антител. Поздние стадии у всех трёх путей активации системы комплемента одинаковы и включают образование мембраноатакующего комплекса, который нарушает целостность мембраны клетки-патогена и приводит к её гибели.

Имму́нная систе́ма — система биологических структур и процессов организма, обеспечивающая его защиту от инфекций, токсинов и злокачественных клеток. Для правильной работы иммунной системы необходимо, чтобы она умела распознавать широкий спектр патогенов — от вирусов до многоклеточных паразитов — и отличать их от собственных здоровых тканей организма. У многих видов имеются две подсистемы: врождённая иммунная система и приобретённая (адаптивная) иммунная система. Обе подсистемы используют как гуморальные механизмы, так и клеточные механизмы. К числу древнейших механизмов иммунной системы животных относят фагоцитоз, систему комплемента и антимикробные пептиды. Адаптивная иммунная система в ходе эволюции появилась у челюстноротых позвоночных животных. Один из важнейших механизмов адаптивной иммунной системы — иммунологическая память, благодаря которой организм развивает более сильный иммунный ответ на патоген после первой встречи с ним. Основу вакцинации составляет именно иммунологическая память.
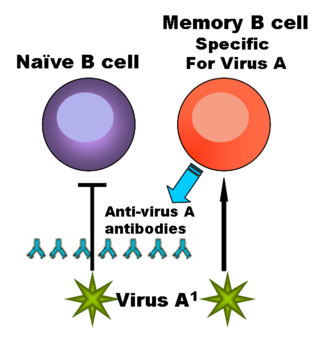
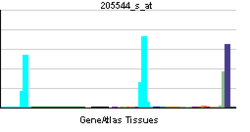
— функциональный тип лимфоцитов, играющих важную роль в обеспечении гуморального иммунитета. При контакте с антигеном или стимуляции со стороны T-клеток некоторые B-лимфоциты трансформируются в плазматические клетки, способные к продукции антител. Другие активированные B-лимфоциты превращаются в B-клетки памяти. Помимо продукции антител, В-клетки выполняют множество других функций: выступают в качестве антигенпрезентирующих клеток, продуцируют цитокины и экзосомы.

C-реактивный белок — белок плазмы крови, относящийся к группе белков острой фазы, концентрация которых повышается при воспалении. Играет защитную роль, связывая бактериальный полисахарид Streptococcus pneumoniae. С-реактивный белок используется в клинической диагностике наряду с СОЭ как индикатор воспаления.

Презентация антиге́на — процесс предъявления T-лимфоциту фрагмента антигена с целью запуска T-клеточного ответа. Поскольку T-клетки не распознают нативные антигены, то нативные антигены предварительно фрагментируются антигенпрезентирующей клеткой, и фрагменты выставляются на её поверхности в связанном с главным комплексом гистосовместимости (MHC) виде, чтобы они могли быть распознаны T-клеточными рецепторами. В случае вирусной или бактериальной инфекции антигенпрезентирующая клетка выставляет на своей поверхности эндогенные или экзогенные пептидные фрагменты, полученные от исходного антигена, в виде комплекса с MHC. Существуют два класса молекул MHC, которые различаются происхождением фрагментов антигена, которые с ними связываются: молекулы MHC I класса (MHC-I) связывают пептидные фрагменты, происходящие из цитозоля клетки, а молекулы MHC II класса (MHC-II) связывают фрагменты экзогенного происхождения, которые появились в результате эндоцитоза исходного антигена и его последующего расщепления. Каждая T-клетка способна распознавать от нескольких десятков до нескольких сотен фрагментов одного и того же антигена, хотя на поверхности антигенпрезентирующей клетки могут быть экспонированы тысячи других пептидных фрагментов, поскольку одна и та же молекула MHC способна связывать самые разнообразные пептиды.
Фолликулярные дендритные клетки (FDC)-это клетки иммунной системы, которые находятся в первичных и вторичных лимфатических фолликулах областей B-клеток лимфоидной ткани. В отличие от дендритных клеток (DC), FDC не происходят от костного мозга гемопоэтических стволовых клеток, а имеют Мезенхима происхождение. Возможные функции FDC включают: организацию клеток и микроархитектуры Лимфатическая система, захват антигена для поддержки B-клеток, содействие удалению мусора из герминальных центров и защиту от автоиммунных заболеваний. Заболевания, в которых могут участвовать FDC, включают первичные опухоли FDC, Воспаление, развитие Вирус иммунодефицита человека инфекции и невроинвазивный Почесуха овец.

Mac-1, или CR3 — мембранный белок, гетеродимерный интегрин подсемейства β2-интегринов, состоящий из альфа цепи αM (CD11b) и бета цепи β2 (CD18). Рецептор комплемента лейкоцитов, играет роль в системе врождённого иммунитета.

TNFRSF17 — мембранный белок, рецептор из надсемейства рецепторов фактора некроза опухоли. Продукт гена человека TNFRSF17. Играет роль в регуляции B-лимфоцитов.

CD19, или B-лимфоцитарный антиген CD19 — белок, ко-рецептор, расположенный на поверхности B-лимфоцитов. Продукт гена человека CD19.

CD22, или рецептор B-клеток CD22 — мембранный белок-рецептор В-лимфоцитов, который у человека кодируется геном CD22. CD22 синтезируется В-клетками, начиная со стадии про-В-клеток, на этом этапе белок находится в цитоплазме. На стадии пре-В-клеток CD22 перемещается в клеточную мембрану и сохраняется на поверхности активированных В-клеток и В-клеток памяти, исчезая только в плазматических клетках. CD22 является ингибиторным ко-рецептором В-клеточного рецептора: он понижает его чувствительность и предотвращает оверстимуляцию клетки антигеном. Этот рецептор является мишенью для некоторых лекарственных препаратов против злокачественных заболеваний крови, которые сейчас проходят клинические испытания.

CCR8 — рецептор β-хемокинов млекопитающих класса интегральных мембранных белков. CCR8 является рецептором для хемокина CCL1. Продукт гена CCR8.

CD81 — мембранный белок из надсемейства тетраспанинов, продукт гена человека TSPAN28.

Фактор ускорения распада комплемента, или CD55 — мембранный белок, ингибитор системы комплемента. Продукт гена человека CD55.

CD23, или Fc-эпсилон-RII (FcεRII), — белок, низкоафинный рецептор иммуноглобулина IgE, изотипа иммуноглобулинов, участвующих в защите от паразитарных инфекций и отвечающих за многие аллергические реакции. В отличие от многих иммуноглобулиновых рецепторов CD23 является лектином типа C. CD23 обнаружен на зрелых B-лимфоцитах, активированных макрофагах, эозинофилах, фолликулярных дендритных клетках и тромбоцитах.

CXCR5 — рецептор хемокинов, продукт гена CXCR5. Относится к семейству хемокиновых рецепторов CXC суперсемейства рецепторов, сопряжённых с G-белком. Обеспечивает миграцию B-лимфоцитов в зону B-клеточных фолликул лимфатических узлов. Рецептор хемокина CXCL13 (BLC). Может участвовать в дифференцировке B-лимфоцитов.

CD3e — трансмембранный белок семейства гликопротеинов CD3, локализованных на поверхности T-лимфоцитов. Продукт гена человека CD3E, расположен на 11-й хромосоме.

Класси́ческий пу́ть актива́ции систе́мы комплеме́нта — один из трёх путей активации системы комплемента, наряду с альтернативным путём и лектиновым путём. Классический путь запускают комплексы антиген-антитело, причём антитела должны принадлежать к классу IgG или IgM. После активации происходит сборка белкового комплекса C3-конвертазы (C4b2b), который разрезает белок комплемента C3. Один из фрагментов C3, C3b, связывается с C3-конвертазой, в результате чего образуется C5-конвертаза (C4b2b3b). C5-конвертаза разрезает компонент комплемента 5, после чего образовавшиеся белковые фрагменты привлекают фагоциты в очаг инфекции и способствуют поглощению клеток патогенов. C5-конвертаза также запускает финальные стадии каскада комплемента, которые завершаются формированием мембраноатакующего комплекса. Он формирует поры в мембране клетки микроорганизма, вызывая её лизис и гибель. Помимо микробных клеток, классический путь комплемента могут запускать апоптотические тельца и некротизированные клетки.
Альтернати́вный путь актива́ции систе́мы комплеме́нта — один из трёх путей активации системы комплемента, наряду с классическим путём и лектиновым путём. В отличие от классического пути, альтернативный путь системы комплемента не требует участия антител, поэтому его относят к гуморальным механизмам врождённого иммунитета.

Рецептор компонента комплемента 5a, тип 1 — мембранный белок семейства рецепторов, сопряжённых с G-белком, рецептор к компоненту комплемента анафилотоксину C5a. Продукт гена человека C5AR1. C5AR1 модулирует воспалительный ответ, а также ассоциирован с ожирением, развитием и онкогенезом.