Органи́ческая хи́мия — раздел химии, изучающий структуру, свойства и методы синтеза соединений углерода с другими химическими элементами, относящихся к органическим соединениям. Первоначальное значение термина органическая химия подразумевало изучение только соединений углерода растительного и животного происхождения. По этой причине ряд углеродсодержащих соединений традиционно не относят к органическим, а рассматривают как неорганические соединения. Условно можно считать, что структурным прототипом органических соединений являются углеводороды.

Прото́н — одна из трёх элементарных частиц, из которых построено обычное вещество. Протоны входят в состав атомных ядер; порядковый номер химического элемента в таблице Менделеева равен количеству протонов в его ядре.

Фотоси́нтез — сложный химический процесс преобразования энергии видимого света в энергию химических связей органических веществ при участии фотосинтетических пигментов.

В биологии активный центр — это область фермента, где молекулы субстрата связываются и подвергаются химической реакции. Активный центр состоит из аминокислотных остатков, которые образуют временные связи с субстратом, и остатков, которые катализируют реакцию этого субстрата. Хотя активный центр занимает только ~ 10-20 % от объёма фермента он является наиболее важной частью, поскольку он непосредственно катализирует химическую реакцию. Обычно активный центр состоит из трех-четырех аминокислот, в то время как другие аминокислоты в белке необходимы для поддержания его третичной структуры.

Свободные радикалы в химии — частицы, содержащие один или несколько неспаренных электронов на внешней электронной оболочке. Свободные радикалы бывают твёрдыми, жидкими и газообразными веществами и могут существовать от очень короткого до очень долгого времени. Радикалы могут быть не только нейтральными, но и ионными, а также иметь более одного неспаренного электрона. Свободные радикалы обладают парамагнитными свойствами и являются очень реакционноспособными частицами.
Дыхательная цепь переноса электронов, также электрон-транспортная цепь (сокр. ЭТЦ, англ. ETC, Electron transport chain) — система трансмембранных белков и переносчиков электронов, необходимых для поддержания энергетического баланса. ЭТЦ поддерживает баланс за счёт переноса электронов и протонов из НАД∙Н и ФАДН2 в акцептор электронов. В случае аэробного дыхания акцептором может быть молекулярный кислород (О2). В случае анаэробного дыхания акцептором могут быть NO3−, NO2−, Fe3+, фумарат, диметилсульфоксид, сера, SO42−, CO2 и т. д. ЭТЦ у прокариот локализована в ЦПМ, у эукариот — на внутренней мембране митохондрий. Переносчики электронов расположены в порядке уменьшения сродства к электрону, то есть по своему окислительно-восстановительному потенциалу, где у акцептора самое сильное сродство к электрону. Поэтому транспорт электрона на всём протяжении цепи протекает самопроизвольно с выделением энергии. Выделение энергии в межмембранное пространство при переносе электронов происходит ступенчато, в виде протона (H+). Протоны из межмембранного пространства попадают в протонную помпу, где наводят протонный потенциал. Протонный потенциал преобразуется АТФ-синтазой в энергию химических связей АТФ. Сопряжённая работа ЭТЦ и АТФ-синтазы носит название окислительного фосфорилирования.

Окисли́тельное фосфорили́рование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Хотя различные формы жизни на Земле используют разные питательные вещества, АТФ является универсальным соединением, в котором запасается энергия, необходимая для других метаболических процессов. Почти все аэробные организмы осуществляют окислительное фосфорилирование. Вероятно, широкому распространению этого метаболического пути способствовала его высокая энергетическая эффективность по сравнению с анаэробным брожением.

Цитохром-c-оксида́за (цитохромоксидаза) или цитохром-c-кислород:оксидоредуктаза, также известная как цитохром aa3 и комплекс IV — терминальная оксидаза аэробной дыхательной цепи переноса электронов, которая катализирует перенос электронов с цитохрома с на кислород с образованием воды. Цитохромоксидаза присутствует во внутренней мембране митохондрий всех эукариот, где её принято называть комплекс IV, а также в клеточной мембране многих аэробных бактерий.
Хиноны — полностью сопряжённые циклогексадиеноны и их аннелированные аналоги. Существуют два класса хинонов: пара-хиноны с пара-расположением карбонильных групп (1,4-хиноны) и орто-хиноны с орто-расположением карбонильных групп (1,2-хиноны). Благодаря способности к обратимому восстановлению до двухатомных фенолов некоторые производные пара-хинонов участвуют в процессах биологического окисления в качестве коферментов ряда оксидоредуктаз.

Феофитин — химическое соединение, служащее одним из первых акцепторов электрона в цепи переноса электронов в реакционном центре фотосистемы II (ФСII) у растений, и в реакционном центре (РЦ) пурпурных бактерий. Как в реакционном центре ФСII, так и бактерий (P870), возбуждённые электроны передаются от воды на феофитин, который затем передаёт их хинону (QA). В целом механизмы, функции и задачи молекулы феофитина аналогичны друг другу в обеих транспортных цепях.

Пластохинон — хинон, вовлечённый в цепь переноса электронов световой фазы фотосинтеза. Во время этой фазы пластохинон восстанавливается (принимает два протона (H+) из стромы хлоропласта, и два электрона (e-) из фотосистемы II), формируя пластохинол. Он транспортирует протоны в полость дисков тилакоида, в то время как электроны передаются по электрон-транспортной цепи к белковому комплексу цитохром b6f.
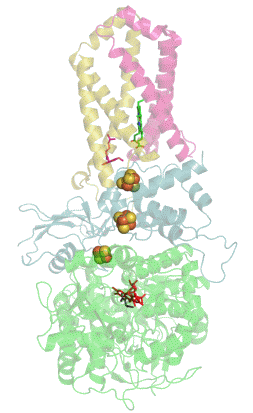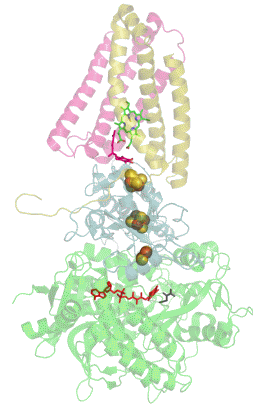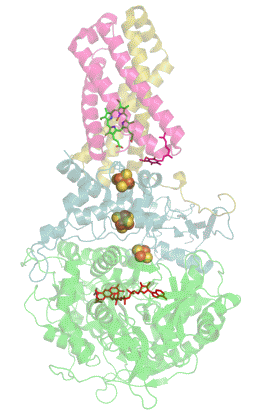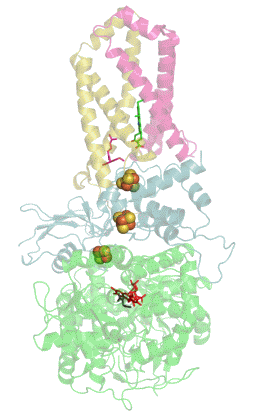
Сукцинатдегидрогеназа или сукцинат-убихинон-оксидорекдуктаза, также известная как комплекс II — белковый комплекс, расположенный во внутренней мембране митохондрий и мембранах многих прокариотических организмов. Одновременно участвует в цикле трикарбоновых кислот и дыхательной цепи переноса электронов.

Фотосисте́ма I, или пластоциани́н-ферредокси́н-оксидоредукта́за — второй функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он принимает электрон от пластоцианина и, поглощая световую энергию, формирует сильный восстановитель П700, способный через цепь переносчиков электронов осуществить восстановление НАДФ+. Таким образом, при участии ФСI синтезируется источник электронов (НАДФН) для последующих реакций восстановления углерода в хлоропластах в цикле Кальвина. Кроме того, ФСI может осуществлять циклический транспорт электронов, сопряжённый с синтезом АТФ, обеспечивая дополнительный синтез АТФ в хлоропластах.

Реакционный центр — комплекс белков, пигментов и других кофакторов, взаимодействие которых обеспечивает реакцию превращения энергии света в химическую при фотосинтезе. Реакционный центр получает энергию или через непосредственное возбуждение одной из своих молекул или через перенос энергии от светособирающих комплексов, что даёт начало цепочке химических реакций, происходящей на связанных белками кофакторах. Эти кофакторы — светопоглощающие молекулы такие как хлорофилл, феофитин и хиноны. Энергия фотона используется для поднятия электрона на более высокий энергетический уровень. Запасённая таким образом свободная энергия идёт на восстановление цепочки акцепторов электрона с более высоким редокс-потенциалом.

Фотосисте́ма II (втора́я фотосисте́ма, фотосисте́ма два, ФСII), или H2O-пластохиноноксидоредуктаза — первый функциональный комплекс электрон-транспортной цепи (ЭТЦ) хлоропластов. Он расположен в мембранах тилакоидов всех растений, водорослей и цианобактерий. Поглощая энергию света в ходе первичных фотохимических реакций, он формирует сильный окислитель — димер хлорофилла a (П680+), который через цепь окислительно-восстановительных реакций способен вызвать окисление воды.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.

НАДН-дегидрогена́зный ко́мплекс, также называемый ко́мплекс I или НАДН-убихино́н-оксидоредукта́за — первый мультибелковый комплекс дыхательной цепи переноса электронов. Множество копий комплекса расположены в мембранах прокариотических организмов, способных к кислородному дыханию и внутренних мембранах митохондрий эукариотических клеток. По отношению к белкам человека комплекс I часто называют НАДН-дегидрогеназой.

Фотофосфорили́рование — процесс синтеза АТФ из АДФ за счёт энергии света. Как и в случае окислительного фосфорилирования, энергия света расходуется на создание протонного градиента на мембране тилакоидов или клеточной мембране бактерии, который затем используется АТФ-синтазой. Фотофосфорилирование — очень древняя форма фотосинтеза, которая есть у всех фототрофных эукариот, бактерий и архей. Различают два типа фосфорилирования — циклическое, сопряжённое с циклическим потоком электронов в электрон-транспортной цепи, и нециклическое, сопряжённое с прямым потоком электронов от H2O к НАДФ+ в случае эукариот или другого донора электрона в случае бактерий, например, H2S. Как разновидность нециклического типа выделяют псевдоциклическое фотофосфорилирование, при котором акцептором электронов служит кислород.

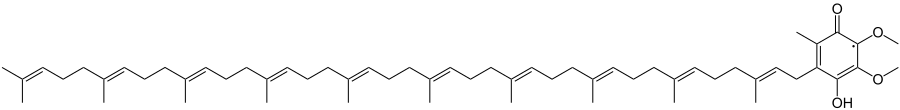
Убихинол — восстановленная форма убихинона. Несёт два дополнительных электрона и два протона. Фактически убихинол можно рассматривать как убихинон, который присоединил к себе молекулу водорода.

1,4-Бе́нзохино́н (хинон, п-бензохинон) — органическое соединение с химической формулой C6H4O2, представитель класса хинонов, один из двух известных изомеров бензохинона. Применяется в производстве гидрохинона, красителей и как дубящее вещество.