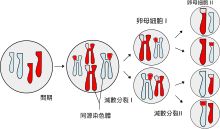
Мейо́з, или редукционное деление — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа. В результате мейоза образуются гаметы, споры и другие зародышевые клетки.

Кроссинго́вер — процесс обмена участками гомологичных хромосом во время конъюгации в профазе первого деления мейоза, которое происходит, например, при образовании гамет или спор. Помимо мейотического, описан также митотический кроссинговер.

Рекомбинация — перераспределение генетического материала путём разрыва и соединения разных молекул, приводящее к появлению новых комбинаций генов или других нуклеотидных последовательностей. В широком смысле слова включает в себя не только рекомбинацию между молекулами ДНК, но и перекомбинацию (сортировку) генетического материала на уровне целых хромосом или ядер, а также обмен плазмидами между клетками.
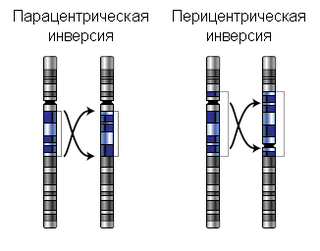
Инве́рсия — хромосомная перестройка, при которой происходит поворот участка хромосомы на 180°. Инверсии являются сбалансированными внутрихромосомными перестройками. Различают парацентрические и перицентрические инверсии. Инверсии играют роль в эволюционном процессе, видообразовании и в нарушениях фертильности.
Парасексуа́льный проце́сс, или парасексуа́льный цикл, — аналог полового процесса, свойственный грибам и одноклеточным организмам. При парасексуальном процессе происходит слияние вегетативных клеток, содержащих генетически разнородные ядра, возникновение гетерозиготных диплоидов за счёт слияния ядер в гетерокарионах и последующего митотического расщепления диплоидов, приводящее к появлению гаплоидных или диплоидных рекомбинантов.
Псевдоаутосо́мные о́бласти — гомологичные участки половых хромосом различного типа; у млекопитающих они, соответственно, находятся на X-хромосоме и Y-хромосоме. Все гены, расположенные в этих областях, есть у обоих полов и наследуются так же, как и любые аутосомные гены, отсюда и название областей.
Хромосомные перестройки — тип мутаций, которые изменяют структуру хромосом. Классифицируют следующие виды хромосомных перестроек: делеции, инверсии, дупликации, транслокации, а также дицентрические и кольцевые хромосомы. Известны также изохромосомы, несущие два одинаковых плеча. Если перестройка изменяет структуру одной хромосомы, то такую перестройку называют внутрихромосомной, если же двух разных, то межхромосомной. Хромосомные перестройки подразделяют также на сбалансированные и несбалансированные. Сбалансированные перестройки не приводят к потере или добавлению генетического материала при формировании, поэтому их носители, как правило, фенотипически нормальны. Несбалансированные перестройки меняют дозовое соотношение генов, и, как правило, их носительство сопряжено с существенными отклонениями от нормы.
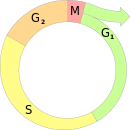
Сестринский хроматидный обмен (СХО) — это обмен участками между сестринскими хроматидами одной хромосомы. Процесс обмена происходит во время S-фазы клеточного цикла путём гомологичной рекомбинации между сестринским хроматидами, генетическая информация при этом остаётся неизменной.

Хромоме́ры — утолщённые, плотно спирализованные участки нитей ДНК (хромонем), из которых состоит хромосома.

Когези́н — это мультибелковый комплекс, который регулирует процесс разделения сестринских хроматид в ходе деления клетки.

Хиа́зма — в генетике точка, в которой две гомологичные несестринские хроматиды обмениваются генетическим материалом в ходе кроссинговера в течение мейоза. Хиазмы становятся видны в фазе диплотены профазы I мейоза, но сам кроссинговер происходит в предыдущей фазе — пахитене. Когда каждая тетрада, состоящая из двух пар сестринских хроматид, начинает распадаться, хиазмы остаются единственными точками контакта.

Бивале́нт — в генетике пара гомологичных хромосом, связывающихся друг с другом во время мейоза посредством специального комплекса после удвоения хромосом. В ходе мейоза происходит синапсис — процесс формирования бивалентов. Каждая из входящих в бивалент хромосом у большинства организмов уже удвоена и состоит из двух хроматид.
Митоти́ческий кроссинго́вер — тип генетической рекомбинации, который может проходить в соматических клетках при митотических делениях как у организмов, обладающих полом, так и бесполых организмов. В случае бесполых организмов митотическая рекомбинация является единственным ключом к пониманию сцепления генов, так как у таких организмов это единственный способ генетической рекомбинации. Кроме того, митотическая рекомбинация может привести к мозаичной экспрессии рецессивных признаков у гетерозиготной особи. Такая экспрессия имеет важное значение в онкогенезе, она также позволяет изучать летальные рецессивные мутации.
Синаптоне́мный ко́мплекс (СК) — белковый комплекс, формирующийся между гомологичными хромосомами в ходе мейоза. Синаптонемный комплекс предположительно является связующим звеном между хромосомами во время спаривания (синапсиса).

Пахитена, или пахинема — это одна из стадий профазы первого деления мейоза, называемая также стадией толстых нитей, во время которой завершается формирование синаптонемного комплекса между гомологичными хромосомами, и происходит кроссинговер. Пахитене предшествуют такие стадии профазы I мейоза, как лептотена и зиготена. После неё следует диплотена и диакинез.

Структу́ра Холлиде́я — структура из четырёх цепей нуклеиновых кислот, соединённых друг с другом водородными связями с образованием четырёх двуцепочечных ветвей. Эти ветви могут принимать несколько различных конформаций в зависимости от концентрации солей в окружающем буферном растворе и последовательности нуклеотидов, располагающихся в непосредственной близости от точки соединения. Структура названа в честь английского молекулярного биолога Робина Холлидея, который предположил её существование в 1964 году.

SMC5 или белок структурной поддержки хромосом номер пять — это белок, который у человека кодируется геном SMC5.

Конденсины — большие белковые комплексы, которые играют главную роль в расхождении хромосом во время митоза и мейоза.

Гомологи́чная рекомбина́ция, или о́бщая рекомбина́ция, — тип генетической рекомбинации, во время которой происходит обмен нуклеотидными последовательностями между двумя похожими или идентичными хромосомами. Это наиболее широко используемый клетками способ устранения двух- или однонитевых повреждений ДНК. Гомологичная рекомбинация также создаёт разнообразие комбинаций генов во время мейоза, обеспечивающих высокий уровень наследственной изменчивости, что, в свою очередь, позволяет популяции лучше адаптироваться в ходе эволюции. Различные штаммы и виды бактерий и вирусов используют гомологичную рекомбинацию в процессе горизонтального переноса генов.
Балансёрная хромосома — элемент генетического инструментария, специально модифицированная хромосома, используемая для поддержания некоторого гена в искусственной популяции в гетерозиготном состоянии и быстрого поиска подобных особей. Также используется для компенсации повреждённого гена и предотвращения рекомбинации (кроссинговера) между гомологичными хромосомами в мейозе. Наиболее часто используется в генетике Drosophila melanogaster, что позволяет поддерживать в популяции мутацию в состоянии гетерозиготы без постоянного контроля её наличия; также может использоваться для мышей. Балансерная хромосома имеет три важных свойства: она подавляет рекомбинацию с гомологичной хромосомой, несёт в себе доминантные маркеры, негативно влияет на репродуктивную функцию в состоянии гомозиготы.