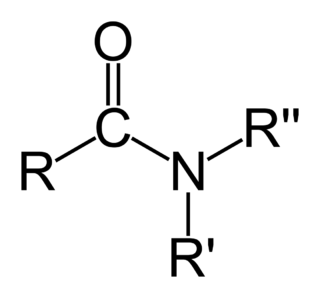
Ами́ды — производные кислородсодержащих кислот, в которых гидроксильная группа кислотного остатка заменена аминогруппой. Амиды также можно рассматривать как ацилпроизводные аминов. Соединения с одним, двумя или тремя ацильными заместителями у атома азота называются первичными, вторичными и третичными амидами соответственно. Вторичные амиды также называют имидами.

Липопротеи́ны (липопротеиды) — класс сложных белков, простетическая группа которых представлена каким-либо липидом. Так, в составе липопротеинов могут быть свободные жирные кислоты, нейтральные жиры, фосфолипиды, холестериды.

Фосфолипи́ды — сложные липиды, представляют собой сложные эфиры многоатомных спиртов и высших жирных кислот. Содержат остаток фосфорной кислоты и соединённую с ней добавочную группу атомов различной химической природы.
Лигаза — фермент, катализирующий соединение двух молекул с образованием новой химической связи — лигирование. При этом обычно происходит отщепление (гидролиз) небольшой химической группы от одной из молекул.

Трегалóза или микóза — органическое соединение, углевод из группы невосстанавливающих дисахаридов. В природной трегалозе 2 остатка D-глюкозы связаны α-1,1-гликозидной связью.
Нуклеазы — большая группа ферментов, гидролизующих фосфодиэфирную связь между субъединицами нуклеиновых кислот. Различают несколько типов нуклеаз в зависимости от их специфичности: экзонуклеазы и эндонуклеазы, рибонуклеазы и дезоксирибонуклеазы, рестриктазы и некоторые другие. Рестриктазы занимают важное положение в прикладной молекулярной биологии.

Фосфодиэстеразы — это группа ферментов, гидролизующих фосфодиэфирную связь. В широком смысле к ним относятся ДНКазы, РНКазы, цАМФ-фосфодиэстеразы, цГМФ-фосфодиэстеразы, фосфолипаза C и фосфолипаза D. Иногда под этим термином подразумевают более узкую группу ферментов, участвующих в регуляции сигнальных путей, то есть в первую очередь цАМФ-фосфодиэстеразы и цГМФ-фосфодиэстеразы.
Фосфолипазы — группа ферментов, которые катализируют процесс гидролиза фосфолипидов. В зависимости от положения гидролизуемой связи в фосфолипиде различают 4 основных класса фосфолипаз: A, B, C и D.

Фосфолипаза A1 — гидролитический фермент, один из типов фосфолипазы, катализирующей процесс элиминирования ацильной цепи фосфолипида в положении sn-1. При действии фосфолипазы А1 на фосфолипид образуется 1-лизофосфолипид и жирная кислота. Фосфолипаза А1 является активным компонентом змеинового яда гемолитического действия.

Фосфолипаза A2 — тип фосфолипазы, гидролизующей ацильную цепь фосфолипида в sn-2 положении. При действии фосфолипазы А2 на фосфолипид образуется 2-лизофосфолипид и жирная кислота. В sn-2 положении фосфолипида могут находиться различные жирные кислоты, включая арахидоновую и эйкозапентаеновую кислоты.
Фосфолипаза B — тип фосфолипазы, обладающей активностями как фосфолипазы А1 так и А2, то есть способной гидролизовать ацильную цепь фосфолипида в положениях sn-1 и sn-2. Как правило фосфолипаза В действует на лизолецитин (лизофосфатидилхолин), образующийся в результате действия фосфолипазы А1 на лецитин (фосфатидилхолин).

Фосфолипаза C — фосфолипаза, гидролизующая фосфодиэфирную связь между глицериновым остатком фосфолипида и полярной фосфатной группой. Относится к фосфодиэстеразам, как и фосфолипаза D. Фосфолипаза С является ключевым ферментом метаболизма фосфатидилинозитола и липидных сигнальных путей.
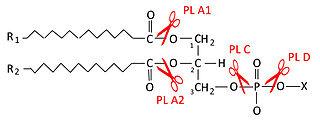
Фосфолипаза D — фосфолипаза, гидролизующая фосфодиэфирную связь между остатком фосфатидной кислоты фосфолипида и головной группой полярной части. Относится к фосфодиэстеразам также как и фосфолипаза C. Существует 2 изоформы этой фосфолипазы D1 и D2. Обе изоформы специфически расщепляют фосфатидилхолин на фосфатидную кислоту и холин, высвобождая последний в цитоплазму. Изоформы имеют различную внутриклеточную локализацию. Фосфолипаза D1 расположена в аппарате Гольджи, эндосомах, лизосомах и в секреторных гранулах, тогда как фосфолипаза D2 локализуется в липидных рафтах плазматической мембраны.
Экзонуклеазы — белки из группы нуклеаз, отщепляющие концевые мононуклеотиды от полинуклеотидной цепи путём гидролиза фосфодиэфирных связей между нуклеотидами.

Рибонуклеотид представляет собой нуклеотид, содержащий рибозу в качестве пентозного компонента. Считается молекулярным предшественником нуклеиновых кислот. Нуклеотиды являются основными строительными блоками ДНК и РНК. Сами рибонуклеотиды являются основными мономерными строительными блоками для РНК. Дезоксирибонуклеотиды, образующиеся в результате восстановления рибонуклеотидов ферментом рибонуклеотидредуктазой (RNR), являются важными строительными блоками ДНК. Существует несколько различий между дезоксирибонуклеотидами ДНК и рибонуклеотидами РНК. Последовательные нуклеотиды связаны друг с другом с помощью фосфодиэфирных связей.

Инозитолтрифосфат (IP3) — это водорастворимый вторичный посредник. IP3 образуется в результате распада мембранных фосфолипидов под действием фермента фосфолипазы С. Инозитолтрифосфат вместе с диацилглицерином принимает участие в передаче сигнала в клетке.
Фактор активации тромбоцитов — сильный фосфолипидный медиатор воспаления. Синтезируется многими типами клеток: нейтрофилами, базофилами, тромбоцитами и эндотелиальными клетками. Участвует в воспалении, агрегации тромбоцитов, играет роль в патогенезе анафилактического шока.

Лейкотрие́ны — органические соединения, группа липидных высокоактивных веществ, образующаяся в организме из арахидоновой кислоты, содержащей 20-членную углеродную цепь. Подкласс лейкотриенов, вместе с простаноидами, входит в класс эйкозаноидов. Один из основных эффектов лейкотриенов — бронхоспазм — лежит в основе патогенеза бронхиальной астмы.
Плазмалогены — фосфолипиды, у которых в первом положении глицерина находится не жирная кислота, а остаток спирта с длинной алифатической цепью, связанный простой эфирной связью.
Липидный обмен, или метаболизм липидов — сложный биохимический и физиологический процесс, происходящий в некоторых клетках живых организмов.