
Пласти́ды — полуавтономные органеллы высших растений, водорослей и некоторых фотосинтезирующих простейших. Пластиды имеют от двух до четырёх мембран, собственный геном и белоксинтезирующий аппарат.
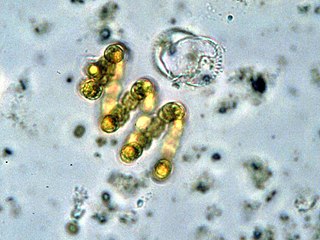
Цианобакте́рии, или сине-зелёные во́доросли, или циане́и — отдел крупных грамотрицательных бактерий, способных к фотосинтезу, сопровождающемуся выделением кислорода.
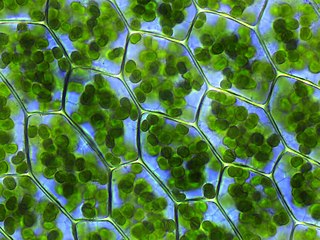
Хлоропла́сты — зелёные пластиды, которые встречаются в клетках фотосинтезирующих эукариот. С их помощью происходит фотосинтез. Хлоропласты содержат хлорофилл. У зелёных растений являются двумембранными органеллами. Под двойной мембраной имеются тилакоиды. Тилакоиды высших растений группируются в граны, которые представляют собой стопки сплюснутых и тесно прижатых друг к другу тилакоидов, имеющих форму дисков. Соединяются граны с помощью ламелл. Пространство между оболочкой хлоропласта и тилакоидами называется стромой. В строме содержатся хлоропластные молекулы РНК, пластидная ДНК, рибосомы, крахмальные зёрна, а также ферменты цикла Кальвина.

Зелёные во́доросли — группа низших растений. В современной систематике эта группа имеет ранг отдела, включающего одноклеточные и колониальные планктонные водоросли, одноклеточные и многоклеточные формы бентосных водорослей. Здесь встречаются все морфологические типы слоевища, кроме ризоподиальных одноклеточных и крупных многоклеточных форм со сложным строением. Многие нитчатые зелёные водоросли крепятся к субстрату только на ранних стадиях развития, затем они становятся свободноживущими, формируя маты или шары.

Эвгленовые — класс протистов, объединяющий около 1000 одноклеточных, реже колониальных видов. Эвгленовые обитают в пресных водоёмах по всему миру. Небольшое число представителей приурочены исключительно к солоноватым и морским водам — роды Eutreptia, Eutreptiella, Klebsina, отдельные виды других родов, где они встречаются в планктоне, зарослях прибрежных водорослей и интерстициали песчаных пляжей.

Динофлагелля́ты, или динофи́товые во́доросли, или динофи́ты, или перидине́и, или па́нцирные жгутиконо́сцы — крупная группа протистов из надтипа альвеолят (Alveolata), которой традиционно присваивают ранг типа. Известно около 4000 ископаемых и более 2500 современных видов, из которых 90 % обитает в морях, остальные — в пресных водах. Около половины представителей — свободноживущие фотосинтезирующие организмы, однако известны и бесцветные гетеротрофные формы, и паразитические динофлагелляты. Некоторые виды являются симбионтами коралловых полипов и двустворчатых моллюсков.

Жёлто-зелёные водоросли, в литературе встречается также вариант Жёлтозелёные водоросли, или Разножгутиковые водоросли, или Трибофициевые — класс водорослей, хлоропласты которых окрашены в жёлто-зелёный или жёлтый цвет. Представители — одноклеточные, колониальные и многоклеточные, преимущественно пресноводные организмы. Аналогично золотистым водорослям, в основу деления жёлто-зелёных на классы положено разнообразие морфологической организации таллома.

Бурые водоросли (лат. Phaeophyceae) — класс отдела охрофитовых водорослей. В жизненном цикле всех представителей присутствуют многоклеточные стадии. Бурые водоросли в хроматофорах содержат бурый пигмент фукоксантин (C40H56O6), который маскирует остальные пигменты.

Печёночные мхи, или Печёночники, или Маршанциевые мхи , — отдел моховидных растений со слабо развитой протонемой. Распространены по всему земному шару. Число видов — от шести до восьми тысяч.

Го́ниум — род колониальных зелёных водорослей из семейства Гониевых (Goniaceae). Это наиболее просто устроенный представитель семейства.

Жгутиковые — жизненная форма протистов. Используют жгутики для локомоции или создания токов воды, приносящих пищу. Среди жгутиконосцев много как свободноживущих форм, так и паразитов и симбионтов животных. Среди них есть одноклеточные моноэнергидные и полиэнергидные формы, а также колониальные и многоклеточные (Volvox) формы. В целом для жгутиконосцев характерна тенденция к мелким размерам клеток и осмотрофному питанию, хотя среди них встречаются также очень крупные фаготрофные формы.

Гаптофитовые водоросли, Гаптофиты, Примнезиофитовые водоросли, или Примнезиофиты — тип эукариот неясного систематического положения. Включает два класса — Павловофициевые (Pavlovophyceae) и Примнезиофициевые (Prymnesiophyceae).
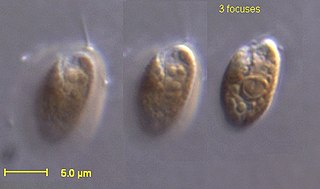
Криптофи́товые во́доросли, или криптомона́ды, или криптофи́ты , — группа одноклеточных эукариотических фотосинтезирующих организмов, включающая около 165 видов, которой традиционно присваивают ранг типа. Почти все криптофитовые имеют монадную форму с дорсовентральным строением, несут два неравных жгутика. Покровы клетки представлены перипластом, имеются стрекательные структуры (эжектосомы). Хлоропласты окружены четырьмя мембранами и содержат редуцированное ядро — нуклеоморф. Основные фотосинтетические пигменты — хлорофиллы a, c2, а также каротиноиды и фикобилины. Митоз обычно открытый, без центриолей, размножение в основном — вегетативное.

Листосте́бельные мхи, или бри́евые мхи, или брио́псиды — класс мхов. В отличие от других моховидных, тело гаметофита листостебельных мхов расчленено на стебель и листья.

Сфагновые мхи — класс растений отдела Моховидные (Bryophyta), ранее этот таксон рассматривали в ранге подкласса (Sphagnidae), используя в качестве русских названий названия Сфагниды либо Сфагновые мхи.

Глаукофи́товые во́доросли, или глаукофи́ты, или глаукоцистофи́ты — небольшой древний отдел одноклеточных водорослей, включающий восемь родов и 21 вид. Глаукофиты особенно интересны своими уникальными хлоропластами (цианеллами), довольно примитивно устроенными по сравнению с хлоропластами других водорослей: они содержат слой муреина между мембранами и характеризуются множеством других свойств, присущих цианобактериям. Согласно современным представлениям, глаукофитовые водоросли обособились в отдельную группу у самого основания Archaeplastida.

Цитохро́м-b6f-ко́мплекс, или пластохинолпластоцианинредукта́за — мультибелковый комплекс, который осуществляет окисление пластохинолов и восстановление белка пластоцианина, обеспечивая, таким образом, транспорт электронов между реакционными центрами фотосистемы I (ФСI) и фотосистемы II (ФСII). Он восстанавливает маленький водорастворимый белок пластоцианин, который переносит электрон к ФСII. Аналогичную реакцию катализирует цитохром-bc1-комплекс электрон-транспортной цепи митохондрий. Цитохром-b6f-комплекс присутствует в тилакоидной мембране хлоропластов растений, водорослей и цианобактерий. Он функционально объединяет две фотосистемы в единую цепь переноса электронов от воды к НАДФ+, то есть является участником нециклического потока электронов. Кроме того, цитохромный комплекс вовлечён в циклический транспорт электронов, осуществляемый фотосистемой I.

Хлорарахниофи́товые во́доросли — клада морских одноклеточных водорослей, входящая в состав группы церкозоев, которым в последних работах систематиков присваивают ранг класса и типа соответственно. Распространены в морях тропического и умеренного поясов. Для организмов этой группы характерно наличие четырёхмембранных хлоропластов, в перипластидном пространстве которых находится нуклеоморф — редуцированное ядро эндосимбиотического эукариота. Хлорарахниофитовые водоросли — миксотрофы, они содержат хлорофиллы a и b и способны к фаготрофному питанию. Размножаются вегетативным и бесполым путём, у некоторых описан половой процесс. По последним данным, клада содержит 14 видов в 8 родах.
C4-фотосинтез, или цикл Хэтча — Слэка, — путь связывания углерода, характерный для высших растений, первым продуктом которого является четырёхуглеродная щавелевоуксусная кислота, а не трёхуглеродная 3-фосфоглицериновая кислота, как у большинства растений с обычным C3-фотосинтезом.
Родельфис — род хищных одноклеточных растительных организмов, родственный к красным водорослям и выделенный в новый отдел эукариот Rhodelphidia. 2 вида.